Measurement of Dynamic Coupling of Bo Hongab, Soumyadipta Acharyaa, Yixuan Kub, Shangkai Gaob, Nitish V. Thakora aDepartment of Biomedical Engineering, Johns Hopkins University, Baltimore, USA Correspondence: Bo Hong, Department of Biomedical Engineering,
Tsinghua University, 100084, Beijing, China. Abstract. Dynamic coupling between different brain regions has been postulated as one of the underlying neural mechanism of cognitive brain activity. Practical methods for accurately quantifying coupling between different brain regions using scalp EEG are plagued by volume conduction effects and background noise. We propose a new method of measuring transient phase locking between independent components underlying cognitive brain activities. This unique combination of independent component analysis (ICA) and phase locking statistics (PLS) provides a promising technique for investigating the dynamics of neural synchronization between different cortical areas, without the contamination of background coherence. This method was tested on the EEG from a cross-modal working memory experiment and its efficacy in detecting transient phase synchrony was demonstrated. Distinct temporal patterns of phase synchrony between prefrontal and parietal components were observed in cross-modal task and in uni-modal task. Keywords: Phase Locking Statistics; Independent Component Analysis; Synchronization; EEG; working memory 1. IntroductionPhase synchrony of oscillatory activity from different neural ensembles is the putative underlying mechanism of neural interaction and integration, through which neural assemblies are dynamically formed to accomplish perceptual, motor and cognitive functions [Schnitzler and Gross, 2005; Singer, 1999; Varela et al., 2001]. This kind of neural synchronization, typically lasting for several to hundreds of milliseconds, spans multiple scales from adjacent neurons to different cortical lobes. In the case of single unit recording, the estimation of cross-correlation between spike occurrences has been widely used for synchronization measurement [Siapas et al., 2005]. However, estimation of synchronization from the scalp EEG remains a challenge, primarily due to volume conduction effects. The signal recorded by an EEG electrode cannot be directly attributed to the underlying cortical area due to the inhomogeneous conducting structure of the human head. Two separate electrodes may actually record signal components from the same neural populations, resulting in spurious synchrony. The other problem lies in the robust estimation of instantaneous phase across various frequency bands in the presence of background noise. Despite the good temporal resolution of these electrophysiological signals, the broad band oscillatory activity is contaminated by considerable background or intrinsic noise, making it difficult to extract transient phase changes in narrow frequency bands and short time windows. The general procedure for detecting and quantifying phase synchrony (or phase-locking) consists of the following steps. Given two signals, their instantaneous phase (across different frequency bands) is estimated by convolution with a complex wavelet or by the Hilbert transform. With minor differences both approaches seem to be fundamentally equivalent [Le Van Quyen et al., 2001]. The phase differences between the signals usually fluctuate around a constant value across trials. It is therefore necessary to test for synchrony in a statistical sense. |



| IJBEM, 2006, Vol. 8, No. 1 | Page VI/2 |
|
The significance of the phase difference between the two signals, for different time latencies, is tested by comparing the Phase Locking Value (PLV) [Lachaux et al., 1999], entropy or mutual information with those obtained from surrogate data. An elegant way of significance testing, known as the phase locking statistics (PLS)[Lachaux et al., 1999], appears to provide a reliable measurement of transient phase coupling. Significant research has been undertaken to improve the spatial resolution of scalp EEG by spatial filtering. Some of these approaches are based on the geometry of the electrodes and the head model. Techniques like the Laplacian derivation[Nunez et al., 1994], inverse electromagnetic methods [Pascual-Marqui, 1999]etc., bring the power of spatial cortical mapping to the scalp EEG. However, the ill-posed nature of the inverse problem still limits the robustness and accuracy of source localization. Alternatively, another category of methods derive spatial filter weights by analyzing the statistical structure of the multi-channel EEG data. Principal component analysis (PCA), independent component analysis (ICA), common spatial subspace decomposition (CSSD), or common spatial pattern, (CSP)[Ramoser et al., 1999] are among them. ICA has been especially successful in analyzing event related brain dynamics [Delorme and Makeig, 2004; Makeig et al., 2004]. Applied to EEG data, ICA analysis leads to spatial filters that separate the recorded scalp activity into a sum of spatially fixed and temporally maximal independent components. By carefully identifying the different spatial distribution patterns and spectrum, some of the decomposed components can be plausibly assigned to underlying brain activity. This provides an elegant way of investigating the dynamics of cognitive processes. To reveal the dynamic interactions between different brain areas during cognitive processes, we propose a method for the measurement of transient phase synchrony between independent components characterized by distinct spatial patterns. 2. Material and Methods2.1. Cognitive EEG data and its processingOur cognitive EEG data were collected in a tactile-visual cross-modal working memory task [Ohara et al., 2005]. In the cross-modal matching task, every trial started with a 100ms tactile vibration stimulus (S1, high frequency 150Hz and low frequency 80Hz) applied to the subjects left index finger-pad, which was followed by a 1.5s delay. At the end of the delay, a visual stimulus of green or red LED (S2) was presented. The subject was instructed to indicate as quickly as possible whether the frequency of S1 and the color of S2 matched by pressing one of two buttons using right hand finger. The color of S2 was associated with the high or low frequency S1 respectively before the experiment, so that the subject could make the decision of match or not. In the control task (unimodal matching task), the sequence of stimuli was identical to the cross-modal task as described above except that the subject was instructed to take S2 as a go-signal, i.e., ignore the color of S2, and to indicate whether S1 is high or low frequency by pressing one of two buttons. During the task performance, scalp EEG was recorded using a 32-electrode recording system (Neuroscan, TX, USA) with a sample rate of 500Hz and an online band-pass filtering (0.1-100Hz). Horizontal and vertical ocular movement were monitored and recorded by two EOG channels. After rejecting EEG portions with ocular artifacts and other evident noise, the 30-channel continuous EEG from 5 subjects was segmented into 2 second epochs (500ms before S1 and 1500ms after S1). Then the average of 500ms before S1 was subtracted as a baseline from the following 1500ms signal. All 1500ms epochs from one subject were concatenated and submitted to an Infomax-ICA decomposition. Using the method as described in section 2.2, two independent components, mostly representing activity from the prefrontal cortex (PFC) and the posterior parietal cortex (PPC) were identified. Hereafter, we only focus on the analysis of phase synchrony between FPC and PPC activities. 2.2 Independent Cognitive Component ExtractionInstead of directly using EEG traces for phase synchronization measurement, independent components (ICs) were first obtained from multi-trial EEG and then identified based on their spatial and temporal properties. ICA decomposes the multi-channel multi-trial EEG into ICs, which makes it possible to pick up source activities localized at different cortical regions of interest. For a multi-channel EEG measurement x = [x1,x2, xn] on the scalp, ICA finds a de-mixing matrix W which makes components of u = Wx mutually independent. To some degree, the independent component u can be an estimate of unknown source matrix s = [s1,s2, , sn] originating from an n-channel observed signal on the scalp. Since there are many different contrast functions to measure the independence of u, W does not have a unique solution. Here the Infomax-ICA [Bell and Sejnowski, 1995] was adopted which achieved the independence by maximizing the mutual information between input x and output u. |
| IJBEM, 2006, Vol. 8, No. 1 | Page VI/3 |
|
Usually, the number of ICs generated from ICA is the same as the number of EEG channels. To find the IC which represents the underlying activity of a cortical area of interest, a strategy similar to the one described in [Xu et al., 2004] was applied. We first defined a subset of electrodes Q={qk}(Electrodes of Interest, EOI), in which qk is the index number of EEG electrodes that cover the cortical area that we want to investigate. For our study, two sets of EOI were defined for prefrontal and parietal areas respectively. The prefrontal EOI consists of 7 electrodes of the 10-20 international system, viz. FP1, FP2, Fz, F3, F4, F7 and F8. The parietal EOI consists of the 6 electrodes P3, Pz, P4, CP3, CPz and CP4. To consider related signal components as much as possible, our EOI definition was broader than the segmentation suggested by the 10-20 international system. In general, the ICs that have relatively large projection on the predefined EOI will be the representative candidates of the given cortical area. Denoting the ith row jth column element of W-1 (the inverse matrix of W) by w'ij, we have
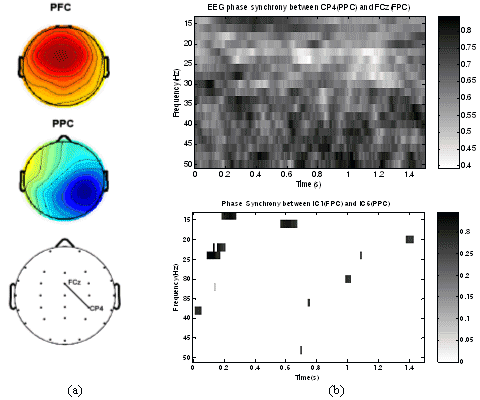
in which the jth column reflects the projection intensity at each electrode of the jth IC uj and can be mapped as a scalp topography (see Fig.1a). We first transformed the spatial pattern matrix W-1 into intensity order matrix M. The value of the element mij in M is the order of the value w'ij in the column vector wj of W-1. For example, if w'ij is the biggest element in column j, then, mij = 1; if w'ij is the second biggest element in column j, then, mij = 2; and so on. Given an electrode subset Q, the representative IC candidates are chosen as uj, where mqkj < T and qk ∈ Q. T is the threshold of admission. For example, setting T to 1 implies that only the ICs with the corresponding electrode of their largest coefficient belonging to the set of EOI can be the candidate; setting T to 2 implies that the largest two are considered; and so on. If necessary, T is increased to ensure at least one candidate can be found. In cases where more than one IC candidates were found, dipole fitting techniques for IC source localization was employed to make further selection of most likely IC (see Fig.2). Usually, a cognitive independent component should have a dipole-like scalp map, i.e., the estimated dipole should have a reasonable position inside the brain and the residual variance of estimation should not be high [Delorme and Makeig, 2004] (10% threshold was set in our case). For our study on cross-modal working memory, the parietal cortex (PPC) and frontal cortex (PFC) were thought to be involved [Fuster, 2003]. Two ICs representing the cognitive activity of PC and FC were selected for each task. Transient phase synchrony analysis was then performed between these two ICs. 2.3. Transient Phase Synchrony Measurement by PLVWe adapted a recently proposed method [Lachaux et al., 1999] for detecting transient phase synchrony in precise frequency bands between different recording sites (independent components in this case). This method uses responses to multiple trials of similar stimuli, and looks for transient phase locking across trials for different time latencies. Given two independent components IC1 and IC2, the procedure calculates a measure of transient phase locking between them. This requires extraction of the instantaneous phase of each signal, across various frequency bands. As a first step, the convolution of the signal (IC1 or IC2) with a complex Gabor wavelet centered at frequency f ( as in Eq.2) was computed.
where σ is the bandwidth parameter, determining the width of the Gaussian window. To enable good time localization, σ was chosen to be 2/f [Lachaux et al., 1999]. The instantaneous phase Ф(t,n) was extracted from this convolution for different time latencies t across trials n (1,2 ..N). For 2 given signals IC1 and IC2 the instantaneous phase difference was then calculated as
The phase locking value (PLV), a measure of the inter-trial variability of this phase difference, was calculated as follows: |
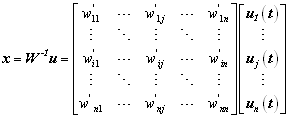
| IJBEM, 2006, Vol. 8, No. 1 | Page VI/4 |
If the phase difference for a given time latency varies very little across trials, then the PLV value (for that value of t) would be close to 1. On the other hand, if there is little inter-trial uniformity, then the PLV value would be close to 0. The next step was to test the statistical significance of the PLV values. This is complicated by the fact that the distribution of the statistics is unknown and uniformity cannot be assumed. We adapted a statistical test based on randomization, which does not require any a priori hypothesis on the signals. For this, 200 surrogate data series were generated by shuffling the trial order for the second signal. These 200 surrogate data sets were used to generate surrogate PLV values. The proportion of surrogate values higher than the original PLV value was designated as the phase locking statistics (PLS) [Lachaux et al., 1999]. We used a significance criteria of 5% (PLS<0.05). This statistical method should be able to detect any significant phase synchrony between the two given signals. 2.4. EEG vs IC phase synchronyTo validate the feasibility and advantage of IC phase synchrony measurement, a comparison between IC phase synchrony and EEG phase synchrony was made. For the two ICs shown in Fig.2a, corresponding two EEG electrodes (FCz and CP4) can be identified at the peak of IC topographies and their scalp position are plotted on Fig.2b. On the time-frequency plane (0-1500ms and 15-50Hz), the PLV were calculated between pair of EEGs and ICs and only those PLV with significance (PLS<0.05) are kept while non-significant PLVs were set to 0. As depicted in the upper inset of Fig.1, the phase synchrony between EEG electrodes is much high across the whole T-F plane, from which it is hard to find a clear pattern. Considering the short distance between these two electrodes, the high synchrony level probably originated from the volume conduction and common noise collected by these two electrodes. For this reason, most of the EEG synchrony and coherence measurement can only be applied on large-scale electrodes instead of local or mesoscale electrodes like FCz and CP4. While after ICA, the background coherence was dramatically eliminated. As shown in the lower inset of Fig. 1b, only some transient phase synchrony was observed.
Figure 1. Comparison of phase synchrony between original EEG channels and independent components. (a) scalp topography of a FPC (or PFC) component (top), a PPC component (middle) and the scalp position of corresponding EEG electrodes (bottom). (b) Phase synchrony of EEG channels(top) on the time-frequency plane compared with that of the independent components(bottom) |
| IJBEM, 2006, Vol. 8, No. 1 | Page VI/5 |
3. Results
|
| IJBEM, 2006, Vol. 8, No. 1 | Page VI/6 |
|
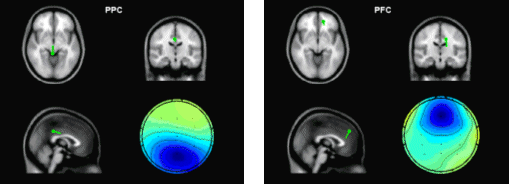
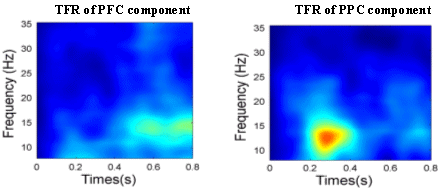
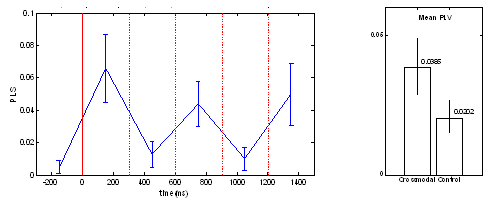
state between PFC and PPC during the period of perceptual memory retention. For the control task, no fixed time pattern found. The position of peak clusters of phase synchrony was variant. In three of the subjects, 2 peaks were observed, while only one cluster of synchrony peak was found in other 2 subjects. The mean PLV averaged over the time span of 0-1500ms is shown in Fig.5b, which indicates that the crossmodal task has a significantly higher level of phase synchrony than that of the unimodal task.
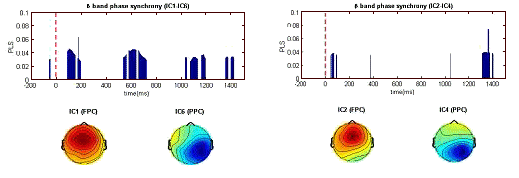
Figure 4. Dynamic pattern of phase synchronyin the β band. Between IC1and IC6 for crossmodal task (left) and between IC2 and IC4 for unimodal task (right).
Figure 5. (a) Dynamic phase synchrony between FPC and PPC components. Three-peak pattern of phase synchrony in the cross-modal task. (b) Mean PLV level (0-1500ms) of crossmodal task and of unimodal task. 4. Discussion and ConclusionsEEG synchronization analysis has the merit of high time resolution and the pitfall of blurred spatial distribution. ICA suppresses the background coherence between scalp EEG channels dramatically, by eliminating the electrode crosstalk caused by volume conduction and separates unrelated noises, while maintaining the same time resolution as EEG. Even though independent components are maximally independent over the whole time range, they may exhibit partial but statistically significant synchronization within specific time/frequency windows [Delorme and Makeig, 2004], through which the transient coupling of neural networks might be revealed. For selecting a physiologically meaningful independent component, spatial or temporal a priori information is needed for the brain activity of interest. The alternative way is to cluster all independent components based on their similarity in spatial distribution (scalp map) and/or time-frequency representation. However, each cluster of independent components has to be assigned to a certain underlying cognitive process by comparing the spatial and/or temporal distribution with a priori ( or predicted) information of that brain process. In [Makeig et al., 2004], Makeig et al. proposed an IC clustering method based on a modified Mahalanobis distance measure of vectors containing map features and spectra of independent component, which ensured a success of revealing the event-related brain dynamics. However, eye inspection was still necessary for some cases and its relative advantage has not been explored. Following this direction, the current framework of transient phase synchrony measurement of ICs might be enhanced by refining the method of automatic IC clustering and optimal selection. Although it is not yet easy to generalize this approach to find new independent brain processes without a priori information, it is still an attractive approach to combine (or validate) dynamics of independent component with other brain imaging methods (e.g. with fMRI, as what was done in [Debener et al., 2005]), to provide further insights about known cognitive process |
| IJBEM, 2006, Vol. 8, No. 1 | Page VI/7 |
AcknowledgementsThis work was supported in part by grant #60205003 from National Natural Science Foundation (China), grant R21NS42690-02 from NIH (USA), and Revolutionary Prosthesis contract from DARPA (USA). The authors would like to thank Dr. S. Ohara and Dr. Y.D. Zhou for providing the data and their constructive comments. 5. ReferencesBell A and Sejnowski T. An information-maximization approach to blind separation and blind deconvolution. Neural Comp 7: 1129-1159, 1995. Delorme A and Makeig S. EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J Neurosci Methods 134: 9-21, 2004. Deneber S, et al. Trial-by-Trial Coupling of Concurrent Electroencephalogram and Functional Magnetic Resonance Imaging Identifies the Dynamics of Performance Monitoring. J Neurosci 25(50):11730-11737, 2005 Fuster J. Cortex and mind: Unifying cognition. New York: Oxford University Press, 2003. Gross J, Schmitz F, Schnitzler I, Kessler K, Shapiro K, Hommel B, and Schnitzler A. Modulation of long-range neural synchrony reflects temporal limitations of visual attention in humans. Proc Nat Acad Sci 101: 13050-13055, 2004. Lachaux J-P, Rodriguez E, Martinerie J, and Varela FJ. Measuring phase synchrony in brain signals. Human Brain Mapping 8: 194-208, 1999. Le Van Quyen M, Foucher J, Lachaux J-P, Rodriguez E, Lutz A, Martinerie J, and Varela FJ. Comparison of Hilbert transform and wavelet methods for the analysis of neuronal synchrony. J Neurosci Methods 111: 83-98, 2001. Makeig S, Delorme A, Westerfield M, Jung T-P, Townsend J, Courchesne E, and Sejnowski TJ. Electroencephalographic Brain Dynamics Following Manually Responded Visual Targets. PLoS Biology 2: 0747-0762, 2004. Nunez P, Silberstein R, Cadusch P, Wijesinghe R, Westdorp A, and Srinivasan R. A theoretical and experimental study of high resolution EEG based on surface Laplacians and cortical imaging. Electroenceph Clinical Neurophysiol:40-57, 1994. Ohara S, Lenz F, and Zhou Y. Sequential neural processes of tactile-visual crossmodal working memory. Neurosci in press, 2005. Pascual-Marqui R. Review of Methods for Solving the EEG Inverse Problem. Int J Bioelectromagnetism 1: 75-86, 1999. Ramoser H, Muller-Gerking J, and Pfurtscheller G. Optimal spatial filtering of single trial EEG during imagined handmovement. IEEE Trans Neural Rehabilitation Eng 8:441-446, 1999. Schnitzler A and Gross J. Normal and pathological oscillatory communication in the brain. Nature Reviews Neurosci 6: 285-296, 2005. Siapas AG, Lubenov EV, and Wilson MA. Prefrontal Phase Locking to Hippocampal Theta Oscillations. Neuron46: 141-151, 2005. Singer W. Neuronal synchrony: a versatile code for the definition of relations? Neuron 24: 49-65, 1999. Varela F, Lachaux J-P, Rodriguez E, and Martinerie J. The brainweb: phase synchronization and large-scale integration. Nature Reviews Neuroscience 2: 229-239, 2001. Xu N, Gao X, Hong B, and al. e. BCI Competition 2003--Data set IIb: enhancing P300 wave detection using ICA-based subspace projections for BCI applications. IEEE Trans Biomed Eng 51: 1067-1072, 2004.
|