 |
International Journal of Bioelectromagnetism Vol. 5, No. 1, pp. 179-180, 2003. |
www.ijbem.org |
|
Co-Localization of Sodium-Transporters
in Glenn Terje Linesac,
Jørn Bodvar Sandeb, Per Grøttumc, Tævje A Strømmeb,
aSimula Research Laboratory, Lysaker, Norway Correspondence: GT Lines, Simula Research Laboratory,
P.O. Box 134, NO-1325 Lysaker, Norway. Abstract. A model
for sodium diffusion under the cell membrane has been developed to study the
interaction between different sodium-transporters. Intracellular concentration
gradients were observed in the simulations, indicating the existence of a
functional sarcolemmal subspace.
Keywords: Diffusion; Sodium Transporters; Transmembrane Current; Fuzzy Space; Simulation 1. Introduction The standard concept of excitation-contraction coupling is that a small current from the L-type Ca2+ channel trigger the ryanodine receptors of the sarcoplasmic reticulum to release a large amount of Ca2+ and hence initiate the contractile apparatus. However, the reverse mode of the Na+/Ca2+ exchanger (NCX) is, under conditions that favor calcium-influx, capable of initiating contractions [Leblanc and Hume, 1990]. These conditions include an increased intracellular sodium concentration, which permits the NCX to go in reverse mode (forward mode is the normal condition, expelling calcium ions in exchange for sodium ions). The intracellular sodium concentration is virtually unchanged during the action potential, but locally beneath the sarcolemma large sodium gradients may build up and thus generate a fuzzy space for subsarcolemmal sodium [Semb and Sejersted, 1996]. If the NCX is localized close to the fast Na+- channel it is possible that the sodium concentration in this subspace is large enough to revert the NCX. In addition, there are Na+/K+-pumps that compete with the NCX for sodium ions. Our goal was to investigate these mechanisms by developing a model that describes the overall interaction between the sodium transporters in sarcolemma taking various geometries and diffusion-coefficients into account. 2. Methods Diffusion of sodium was studied in a small neighbourhood around a single Na+-channel. To reduce the computational load, symmetry in the membrane plane around the sodium channel was assumed. The domain of the diffusion problem is then reduced from a volume to a rectangle. One side will correspond to the cell membrane, the other three sides will be artificial, internal boundaries, see Fig. 1. The governing equation in the interior of the domain is a diffusion equation, derived from conservation of mass and Ficks law. On the boundary at the side most distal to the membrane a fixed concentration is assigned. No-flux boundary conditions are given on the two remaining sides. On the membrane side there are three different types of channels that influence the sodium concentration, one Na+-channel, 10 Na+/K+-pumps and 20 Na+/Ca2+-exchangers. The interchannel distance is 14.7 nm. This corresponds to a measured number of channels of 3, 300 and 1200 per mm2, respectively. The transmembrane currents were calculated using the model of Winslow et al [Winslow et al, 1999]. The diffusion coefficient, k in Fig. 1, was varied between 10 nm2/ms and 10000 nm2/ms. The system was formulated as a finite element model and implemented in Diffpack [Langtangen, 1999].
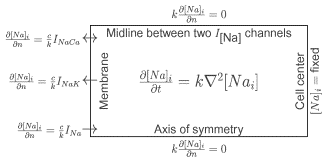 Figure 1. Schematic model with membrane and the 3 ion channel types to the left, boundary conditions and diffusion equation.
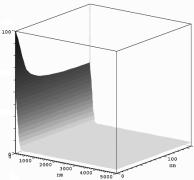 Figure 2. Simulation snapshot at 0.5 ms. The bottom plane corresponds to Fig. 1. The Na+-channel is located in origo.The sodium level is on the ordinate. The diffusion coefficient is 1000 nm2/ms.
3. Results The geometrical resolution of the finite element system was 1 nm around the membrane and gradually coarser into the cell, and about 1 mm at the cell center. The time step was 0.1 ms. Figure 2 shows the sodium concentration over the computational domain 0.5 ms after the onset of stimulus in a simulation with a diffusion coefficient of 1000 nm2/ms. At this point of time the Na+/K+-pump and the NCX show no effect on the system. The sodium concentration is highest around the Na+-channel (in origo) and there is also a distinct layer of elevated concentrations close to the membrane, indicating the existence of a restricted subspace. However, gradients tangential to the membrane surface dissipated very quickly, and the sodium concentration was almost restored to a uniform level right after the action potential upstroke. With higher diffusion coefficients the concentrations gradients disappeared. With lower coefficients the gradients persisted for a longer time. 4. Discussion The computational model was able to reproduce the sodium gradients observed between the fuzzy space and the cytoplasm using realistic geometries, well-established ionic models and sodium diffusion coefficients within the reported range. However, the actual diffusion coefficient is not well established, and values of 10 to 10000 nm2/ms have been reported [Wendt-Gallitelli et al, 1993, Kabakov, 1998]. The simulations indicate that the time course and the magnitude of the sodium concentrations, and thereby the functional significance of the sarcolemmal subspace, depend very much on the precise value of the diffusion coefficient. The simulations have thus helped identify a problem for experimental study. The results further imply that the use of average values for the entire cellular compartment may constitute an erroneous simplification in some modelling studies. References Kabakov AY. Activation of KATP channels by Na/K pump in isolated cardiac myocytes and giant membrane patches. Biophysical Journal, 75:2858-2867, 1998. Langtangen HP. Computational Partial Differential Equations. Numerical Methods and Diffpack Programming. Springer, Berlin, 1999. Leblanc N, Hume JR. Sodium current-induced release of calcium from cardiac sarcoplasmic reticulum. Science, 248:372-376, 1990. Semb SO, Sejersted OM. Fuzzy space and control of Na+, K(+)-pump rate in heart and skeletal muscle. Acta Physiologica Scandinavica, 156:213-225, 1996. Wendt-Gallitelli MF, Voigt T, Isenberg G: Microheterogeneity of subsarcolemmal sodium gradient: electron probe microanalysis in guinea-pig ventricular myocytes. Journal of Physiology, 472:33-44, 1993. Winslow RL, Rice J, Jafri S, Marbán E, ORourke B. Mechanisms of altered excitation-contraction coupling in canine tachycardia-induced heart failure, II Model studies. Circulation Research, 84:571-586, 1999.
© International Society for Bioelectromagnetism
|

