Correspondence: Mark E.
Pflieger Source Signal Imaging 2323 Broadway,
Suite 102 San Diego, CA 92102 USA
E-mail: mep@sourcesignal.com
1. Introduction
Brain dynamics implement brain functions
by organizing brain activity; but we do not directly observe
functions, activity, or dynamics in a functional neuroimaging
experiment. Although the successful performance
of a task manifests brain functions, we observe only the
sensory and motor task variables—not the underlying
brain functions that coordinate, monitor, predict, plan,
and control these variables. We observe shadows
of brain activity via the spatiotemporal signals obtained
from a modality, such as BOLD (blood oxygenation level
dependent) signals from fMRI, or fields/potentials from
M/EEG. However, in order to make inferences about
the underlying brain activity, we must apply biophysical,
physiological, and biostatistical theories to the given
task variables and measured signals. Finally, we
clearly do not directly observe the underlying brain dynamics,
which are the deterministic and/or stochastic laws that
govern brain activity to realize functional requirements.
In all three cases, we must combine the available data
with a body of theory in order to make inferences about
the interesting, underlying unknowns.
Different modalities measure different signals that may
reflect essentially the same underlying brain activity,
dynamics, and functions. Armed with complementary
datasets such as fMRI and M/EEG that have different spatial
and temporal characteristics, the general question is:
Can we make stronger inferences about each of the interesting
unknowns via spatiotemporally improved signal detection
and parameter estimation? Clearly, we will need
some common theoretical machinery in order to make joint,
multimodal inferences.
To begin, let us make an assumption. When different-session
fMRI and M/EEG datasets are available for the same subject
who performed realizations of the same task, let us assume
that the underlying brain dynamics were statistically
equivalent across sessions. Although the brain activities
must have differed in details across sessions, we assume
that the underlying dynamics were substantially equivalent
in the sense that between-session differences were statistically
small compared with other experimentally significant variables.
Based on this testable assumption, our goals are (A) to
design cross-modal analyses of both datasets (fMRI and
M/EEG) that improve, spatially and temporally, the estimation
and detection of the common brain activity, and (B) to
characterize the underlying dynamics. Ultimately,
we seek (C) to understand how the dynamics implement brain
functions in the context of successful task performances.
While keeping in mind the larger context of problem C,
this paper presents an approach for solving problem A
that leads to the threshold of problem B.
1.1 Problem C: Spatiotemporal Integration Functions
In the long term, an ideal analysis of spatiotemporal
imaging data should permit us to make inferences about
brain processes as realizations of brain functions, i.e.,
as solutions to natural problems that engage the brain.
Thus, we ultimately seek methods that can bridge the gap
between (i) raw neuroimaging measurements from various
modalities and (ii) descriptions of underlying brain processes
in terms of the problems they solve. Such descriptions
call for “functional insight” that is attuned
to the “native interests” of the brain regions
under study (e.g., Lettvin et al., 1959).
We can approach this objectively if we express a putative
functional insight as a testable hypothesis that explicitly
formulates the problem that a brain process solves.
Because the analysis we ultimately seek should permit
us to test such functional hypotheses, it follows that
the technical problem of designing imaging algorithms
depends on the scientific problem of formulating
functional theories about the natural problems
solved by the brain. In short, our methods must
match the subject matter. On the other hand, because
the subject matter comprises unresolved functional issues
that our methods should address, we must design the analysis
to match a general theoretical framework that is relatively
neutral, which can accommodate various specific functional
hypotheses. Some viable candidates for such a framework
are: adaptive system theory (Ashby, 1960; Sommerhoff,
1974); schema theory (Arbib et al., 1998); and
information theory used as a language to express functional
hypotheses, e.g., as used by Phillips and Singer (1997)
to formulate the coherent infomax hypothesis of cortical
processing. To our knowledge, these frameworks remain
undeveloped along lines that would render them immediately
useful for the design of imaging analysis algorithms.
However, in spite of this present lack of a suitable general
framework for describing brain function, these basic considerations
nevertheless can guide our current efforts toward the
long-term goal.
Spatiotemporal integration functions play a central role
in solving various problems that engage the brain, which
we classify as extrinsic or intrinsic. “Extrinsic
problems” challenge the brain to dynamically organize
skeletal and visceral behaviors that can achieve co-adaptations
between the organism’s environment and the organism’s
physiology. Two extremes of such co-adaptive processes
are environment-directed and physiology-directed brain
processes. Environment-directed processes adaptively
control environment variables around the organism by coordinating,
in space and time, two streams of brain processes: (a)
motor planning and control of skeletal effector variables,
and (b) sensory monitoring and prediction of the environment
variables. (We use the terms “coordination”,
“planning”, “control”, “monitoring”,
and “prediction” to describe objective forms
of brain function without ascribing subjective content.)
To illustrate, consider Sommerhoff’s (1974) example
of a bird pecking at a grain of seed, the position of
which may be subject to wind perturbations: The planned/controlled
trajectory of the beak must be coordinated with the monitored/predicted
trajectory of the grain. In a formally similar way,
physiology-directed processes modulate the autonomic regulation
of physiological variables in the organism by coordinating,
in space and time, two streams of brain processes: (a)
planning and control of visceral effector variables, and
(b) monitoring and prediction of the physiological variables.
For example, efforts of biomedical engineers to externally
control hemodynamic variables (Isaka and Sebald, 1993;
Palerm et al., 1999) underscore the highly nonlinear
nature of these dynamic physiological control problems.
More generally, co-adaptive processes are bi-directional
and higher-order in the sense that they coordinate environment-directed
(somatic) and physiology-directed (visceral) processes
(Yates and Stocker, 1998) as, for example, when a runner’s
physiology responds to meet the increased demand of climbing
a steep grade. In summary, the requirement for coordination
of multiple brain processes in space and time implies
that spatiotemporal integration functions play a key role
in solving extrinsic problems.
“Intrinsic problems” are self-referential
problems that the brain solves (in the context of solving
extrinsic problems) by coordinating brain architectural
development processes and brain physiological regulation
processes. Thus, two extreme types of processes
that solve intrinsic problems are neurophysiological self-regulation
and neuroarchitectural self-organization. Processes
of the former type modulate the regulation of physiological
variables in the brain itself, and so are special cases
of the physiology-directed processes discussed above.
An example is the regulation of the coupling between brain
hemodynamic variables (such as regional cerebral blood
flow) and neuronal activity variables (such as mean firing
rate). In order to connect the neuronal and the
hemodynamic variables in this example, spatiotemporal
integration must span different temporal and spatial scales.
At the other extreme, architectural self-organization
(which descends from genetic and embryological processes)
implies that the brain “develops itself”,
with each developmental stage setting new constraints
and providing new possibilities for the next stage (Kelso,
1995; Arbib et al., 1998). For example, functional
segregation constrains the dynamics of the adult brain
while simultaneously providing combinatorial possibilities
for functional integration. This modular architecture
is a general solution to an intrinsic “multimodal
integration” problem (Sporns et al., 2000),
which conditions the ongoing solving of particular “dynamic
link” architectural problems that arise in the context
of solving extrinsic problems (von der Malsberg and Singer,
1988).
The concept of neuronal transients (Friston, 1997; 2000)
arises in the context of this intrinsic multimodal integration
problem. Although each specialized area operates
on its own characteristic spatial and temporal scale,
brain areas collectively coordinate their functions, as
necessary. In order to provide coherent output signals
to other brain areas and/or motor systems, each area must
therefore resolve spatial ambiguities and temporal asynchronies
in its input signals as they arrive from other brain areas
and/or sensory modalities. In particular, finite
intervals of integration are required in order to resolve
uncertainties that arise from temporal asynchrony (Pöppel,
1997). This necessitates asynchronous coupling between
neuronal transients in different brain regions, which
in turn requires nonlinear interactions (Friston, 2000).
Others (e.g., Poggio and Reichardt, 1981; Koch, 1999)
have emphasized the importance of nonlinearities, such
as multiplication of two or more variables, for realizing
computational requirements for basic functions, such as
pattern recognition, coordination, and control.
Thus at minimum, a model of the nonlinear, asynchronous
dynamics of neuronal transients must be multilinear (permitting
multiplications) and have transient memory (permitting
temporal interactions). The satisfaction of these
conditions by the deterministic Volterra model (Poggio
and Reichardt, 1981; Friston, 2000) sets the stage for
using nonlinear system identification methods to approach
problem A.
1.2 Problem A: Solution Strategy
Our strategy for solving problem A is as follows.
Task-related M/EEG or fMRI analyses treat the brain as
a dynamic system, where the input variables are sensory
and/or motor task variables, the output variables are
measured M/EEG or fMRI signals, and the analysis yields
estimates of system parameters that maximize the dynamic
coupling between the input and the output variables.
Let us conduct a thought experiment that suspends the
physical impossibility of simultaneous MEG and fMRI:
Suppose that we have an ideal, simultaneous M/EEG and
fMRI dataset. We could perform cross-modal analyses
with such a dataset—call these “fMRI-related
M/EEG” and “M/EEG-related fMRI”—by
using the measurements of one modality as the input variables,
in place of the sensory and/or motor task variables.
Here we suppose that the analysis method can handle continuous
as well as discrete (“event-related”) input
variables. An estimator of M/EEG source current
activity at a given location in the brain, for a given
orientation, has the form of a linear combination of channels.
Using an fMRI BOLD signal at a voxel (or a local average
of voxels), we can design an fMRI-related M/EEG analysis
that “tunes” the linear estimator of source
current activity to maximize the dynamic coupling between
the estimated activity and the BOLD signal. Then
we can use this M/EEG-derived estimate of neuronal activity
to make an M/EEG-related fMRI analysis that estimates
the hemodynamic response to neuronal activity at the voxel
(or local average of voxels). Because task variables
are remote from the hemodynamic system, the “neuronal-derived”
hemodynamic response will differ from the task-derived
hemodynamic response. Consequently, matched filter
signal detection using the neuronal-derived hemodynamic
response may be more powerful for constructing fMRI statistic
images of activated voxels. In summary, fMRI spatially
focuses M/EEG source time series estimates, which we use
to produce an improved estimate of the fMRI hemodynamic
response to neuronal activity and to derive a new fMRI
statistic image. Thus, the strategy is to use the
temporal strengths of M/EEG to enhance the spatial strengths
of fMRI, and vice versa.

Figure 1.
Schematic of the overall strategy for solving
problem A. System models that maximize
the dynamic coupling between task variables
and measured signals are independently identified
for same-subject, same-task, different-session
fMRI and M/EEG datasets. These models
are used to simulate a simultaneous fMRI and
M/EEG dataset. Then cross-modal system
models are identified by maximizing the dynamic
coupling between the simulated fMRI and M/EEG
signals.
To enable this strategy, we must handle the typical situation
where we do not have simultaneous datasets. Our
auxiliary strategy is to use the same dynamic system modeling
methods (that enable the fMRI-related M/EEG and M/EEG-related
fMRI analyses) to make task-related dynamic models, separately
for each modality. Then we can use these dynamic
models to simulate “virtual simultaneous”
fMRI and M/EEG datasets. Figure 1 summarizes the
overall strategy.
It remains to specify the nature of the system model.
Based on the previous section’s considerations,
we require a nonlinear model with transient memory, such
as the Volterra model. Because problem B is concerned
with the underlying dynamics of brain activity, we approach
the question of selecting a dynamic system model in the
context of a formal statement of problem B.
1.3 Problem B: Dynamics and Observables
At a hypothetical and formal level, we can state problem
B in terms of a system of equations that comprise one
state equation (1), which governs the spatiotemporal
dynamics of brain activity, and one observation equation
(2) per modality (Ozaki, 1985; Valdes et al.,
1999):
| |
 |
(1) |
| |
 |
(2) |
where  is the (unknown) state of neuronal
activity at location
is the (unknown) state of neuronal
activity at location  and time
and time  ;
;  is an (unknown) differential operator;
is an (unknown) differential operator;
 is a (known) input vector of control variables;
is a (known) input vector of control variables;
 is an (unknown) deterministic “driving”
function; and
is an (unknown) deterministic “driving”
function; and  is an (unknown) stochastic term.
For the observation equations,
is an (unknown) stochastic term.
For the observation equations,  is an observed measurement we obtain
from neuroimaging modality
is an observed measurement we obtain
from neuroimaging modality  ;
;  is a modality-dependent transfer function
that maps the neuronal state onto the observables via
biophysics, physiology, and instrumentation; and
is a modality-dependent transfer function
that maps the neuronal state onto the observables via
biophysics, physiology, and instrumentation; and  is a noise term. Then given a
set of measurements
is a noise term. Then given a
set of measurements  , control variables
, control variables  , and knowledge of the transfer functions
, and knowledge of the transfer functions
 , we seek to infer first
, we seek to infer first  in both space and time, then ultimately
the dynamics
in both space and time, then ultimately
the dynamics  and
and  (i.e. the specific form of the state
equation itself). When multimodal information is
available, this system of equations implies that our ultimate
goal is to obtain a simultaneous solution, i.e., a joint
inference about the interesting unknowns.
(i.e. the specific form of the state
equation itself). When multimodal information is
available, this system of equations implies that our ultimate
goal is to obtain a simultaneous solution, i.e., a joint
inference about the interesting unknowns.
Simplifications of equation (1) lead to three working
models of the dynamics of task-related neuronal transients:
| |
 |
(3) |
| |
 |
(4) |
| |
 |
(5) |
In all three cases, we expand the driving function  as a Volterra series (see equation
(6)). Thus, the pure Volterra model of equation
(3) directly expands the brain activity as a deterministic
Volterra series of the task variables
as a Volterra series (see equation
(6)). Thus, the pure Volterra model of equation
(3) directly expands the brain activity as a deterministic
Volterra series of the task variables  (plus the stochastic activity).
In the differential-Volterra model of equation
(4), a differential operator
(plus the stochastic activity).
In the differential-Volterra model of equation
(4), a differential operator  characterizes the driven process, which
is a derivation of the neuronal transient activity.
In the self-history Volterra model of equation
(5), we include the brain activity as a variable that
contributes to its own driving function. Treatment
of the stochastic term involves modeling (e.g., autoregressive
modeling) of the task-unrelated brain (and other noise)
processes to account for serially correlated background
activity (Zarahn et al., 1997; Friston, Josephs,
et al., 2000).
characterizes the driven process, which
is a derivation of the neuronal transient activity.
In the self-history Volterra model of equation
(5), we include the brain activity as a variable that
contributes to its own driving function. Treatment
of the stochastic term involves modeling (e.g., autoregressive
modeling) of the task-unrelated brain (and other noise)
processes to account for serially correlated background
activity (Zarahn et al., 1997; Friston, Josephs,
et al., 2000).
Generalizations (4) and (5) may contribute important
theoretical and practical advances over the pure Volterra
model. For example, self-history and nonlinear differential
operators are two ways of modeling deterministic-stochastic
interactions between  and
and  , which may provide a framework for explaining
the observations of Brandt and Jansen (1991) that “deterministic”
visual evoked responses interact with their “stochastic”
prestimulus baselines. Deterministic-stochastic
interactions may also provide an explanatory framework
for well-known (but poorly understood) response variability
phenomena, such as statistically significant single-trial
ERP latency jitter (e.g., Pflieger, 1991). In practice,
both approaches have the potential for more efficient
modeling of the system: Recent self-history may reduce
the history required for task variables; and a few differential
parameters may “leverage” expensive Volterra
kernels.
, which may provide a framework for explaining
the observations of Brandt and Jansen (1991) that “deterministic”
visual evoked responses interact with their “stochastic”
prestimulus baselines. Deterministic-stochastic
interactions may also provide an explanatory framework
for well-known (but poorly understood) response variability
phenomena, such as statistically significant single-trial
ERP latency jitter (e.g., Pflieger, 1991). In practice,
both approaches have the potential for more efficient
modeling of the system: Recent self-history may reduce
the history required for task variables; and a few differential
parameters may “leverage” expensive Volterra
kernels.
1.4 Possibilities for Combined Analysis of fMRI and
M/EEG Datasets
Now we turn specifically to fMRI and M/EEG modalities.
Figure 2 diagrams the relationship between the measured
M/EEG and fMRI signals and the underlying neuronal transient
activity.

Figure 2.
Common underlying sources of M/EEG and fMRI signals.
Neuronal transient activity  (a function of space
(a function of space  and time
and time  ) underlies observed M/EEG signals across
channels
) underlies observed M/EEG signals across
channels  (functions of time) and fMRI BOLD signals
(functions of time) and fMRI BOLD signals
 (a function of space and time).
We assume that the task-related dynamics of neuronal transients
have the form of a differential equation
(a function of space and time).
We assume that the task-related dynamics of neuronal transients
have the form of a differential equation  , where
, where  is a differential operator,
is a differential operator,  is a deterministic driving function
of the task variables
is a deterministic driving function
of the task variables  (functions of time) and self-history,
and
(functions of time) and self-history,
and  represents stochastic influences (a
function of space and time). The transfer functions
represents stochastic influences (a
function of space and time). The transfer functions
 and
and  describe the net effect of biophysical,
physiological, and instrumental factors that convert the
underlying neuronal transient activity to the measured M/EEG
and fMRI signals, respectively. The dashed ellipse
represents a subspace of neuronal transients that can be
sources of M/EEG signals; this subspace is the domain of
the M/EEG transfer function
describe the net effect of biophysical,
physiological, and instrumental factors that convert the
underlying neuronal transient activity to the measured M/EEG
and fMRI signals, respectively. The dashed ellipse
represents a subspace of neuronal transients that can be
sources of M/EEG signals; this subspace is the domain of
the M/EEG transfer function  . Likewise, the dotted ellipse represents
the source domain of the fMRI signals. We assume that
the M/EEG and fMRI source domains overlap, i.e., that there
is a neuronal transient subspace that is the source of both
M/EEG and fMRI BOLD signals. The diamond represents
the neuronal transient subspace sampled by an experiment.
Subspace
. Likewise, the dotted ellipse represents
the source domain of the fMRI signals. We assume that
the M/EEG and fMRI source domains overlap, i.e., that there
is a neuronal transient subspace that is the source of both
M/EEG and fMRI BOLD signals. The diamond represents
the neuronal transient subspace sampled by an experiment.
Subspace  represents those neuronal transients
in the experiment that affect M/EEG, but not fMRI, signals.
Subspace
represents those neuronal transients
in the experiment that affect M/EEG, but not fMRI, signals.
Subspace  represents neuronal transients
that affect fMRI, but not M/EEG, signals. The common
subspace
represents neuronal transients
that affect fMRI, but not M/EEG, signals. The common
subspace  represents neuronal transients
that affect both M/EEG and fMRI signals (whereas subspace
represents neuronal transients
that affect both M/EEG and fMRI signals (whereas subspace
 represents those that affect neither).
The image of the common subspace
represents those that affect neither).
The image of the common subspace  is
is  in the M/EEG signal space, and
in the M/EEG signal space, and
 in the fMRI signal space.
The signal subspaces
in the fMRI signal space.
The signal subspaces  (in M/EEG signal space) and
(in M/EEG signal space) and  (in fMRI signal space) are
images of the disjoint neuronal activity subspaces
(in fMRI signal space) are
images of the disjoint neuronal activity subspaces  and
and  .
.
It is generally believed that postsynaptic neuronal currents
in an open field dendritic arrangement, such as the apical
dendrites of pyramidal cells, form the basis for the scalp
potentials and fields measured as M/EEG (Lorente de Nó,
1947). We obtain the observation equation that relates
these currents and fields from Maxwell's equations, solved
numerically for the volume conductor that represents the
head. This is often referred to as the forward problem.
Volume conduction is linear and, for signal frequencies
less than ~1000 Hz, instantaneous (quasi-static approximation;
Malmivuo and Plonsey, 1995). Forward solution methods
include analytic/spherical (e.g., Arthur and Geselowitz,
1970), boundary element (e.g., Hämäläinen and Sarvas,
1987), and finite element (e.g., Awada and Greenblatt,
1999). However, Helmholtz (1853) showed that there
is no unique solution to the inverse problem of estimating
these neuronal currents from measured extracranial fields.
All such solutions are inherently model-dependent.
The most commonly employed signal in fMRI is blood oxygenation
level dependent (BOLD) contrast: a change in water proton
signals caused by changes in the relative concentration
of venous blood deoxyhemoglobin, which serves as an endogenous
paramagnetic contrast agent (Kwong et al., 1992;
Ogawa et al., 1992). The fMRI observation
equation must incorporate physiological mechanisms that
relate neuronal activity to BOLD signals via local blood
oxygenation, regional blood flow, and regional blood volume—the
theoretical details of which are current topics of research
(Buxton et al., 1998). There are three main
phenomenological components of the (visual) evoked BOLD
response. A small signal decrease (initial dip)
that peaks about 2 s after stimulus onset (not routinely
seen in low fields) results from a transient decrease
of blood oxygenation due to local tissue demands (Menon
et al., 1995). The largest BOLD component,
a positive overshoot that peaks about 5 s, reflects a
decrease in relative deoxyhemoglobin concentration caused
by increased local blood flow in excess of tissue demands.
Finally, a negative undershoot requires up to 30 s to
return to baseline. Timing details of the BOLD signal
are task dependent, and vary across brain regions.
Because the BOLD response to task events is generally
not identical with the response to neuronal activity,
recent efforts (Ogawa et al., 2000) have sought
to relate the BOLD response empirically to neuronal activity
as reflected by EEG. It is unknown whether neural-hemodynamic
decoupling that occurs during normal non-REM sleep (Klingelhofer
et al., 1995) and normal aging (D’Esposito et
al., 1999) may play a wider role during other normal
functions, or perhaps in some brain disorders.
What, then, can we hope to learn about
the underlying neuronal transients by combining information
from M/EEG and fMRI? Of course, we cannot
expect to gain information about subspace  of Figure 2, which represents brain
activity that is invisible to both modalities. Similarly,
because all information we have about subspace
of Figure 2, which represents brain
activity that is invisible to both modalities. Similarly,
because all information we have about subspace  is contained in the M/EEG data,
and all information we have about subspace
is contained in the M/EEG data,
and all information we have about subspace  is contained in the fMRI data,
we cannot expect integrated analyses to cast new light
here either: The most we can hope for is to delineate
the boundaries of these disjoint subspaces. Possibilities
for
is contained in the fMRI data,
we cannot expect integrated analyses to cast new light
here either: The most we can hope for is to delineate
the boundaries of these disjoint subspaces. Possibilities
for  ,
,  , and
, and  include the following: MEG is insensitive
to radial current sources; both EEG and MEG are insensitive
to currents generated in closed field dendritic arrangements
(e.g., basal dendrites of pyramidal cells), and more generally
to unsynchronized local activity (Nunez, 1995); fMRI is
insensitive to small or brief changes in neuronal activity
that do not produce detectable BOLD responses; and neural-hemodynamic
decoupling may occur to some extent in normal and abnormal
brain functioning. For our purposes, it is the common
subspace
include the following: MEG is insensitive
to radial current sources; both EEG and MEG are insensitive
to currents generated in closed field dendritic arrangements
(e.g., basal dendrites of pyramidal cells), and more generally
to unsynchronized local activity (Nunez, 1995); fMRI is
insensitive to small or brief changes in neuronal activity
that do not produce detectable BOLD responses; and neural-hemodynamic
decoupling may occur to some extent in normal and abnormal
brain functioning. For our purposes, it is the common
subspace  , which is visible to both modalities,
that provides the most promising territory for joint analyses.
Here we can reasonably hope that M/EEG and fMRI will provide
complementary information in space and time about the
same underlying brain activity. Thus, two fundamental
objectives of integrated analysis are (a) to identify
the image subspaces
, which is visible to both modalities,
that provides the most promising territory for joint analyses.
Here we can reasonably hope that M/EEG and fMRI will provide
complementary information in space and time about the
same underlying brain activity. Thus, two fundamental
objectives of integrated analysis are (a) to identify
the image subspaces  and
and  in the combined M/EEG and fMRI datasets
that refer to the same underlying brain activity subspace
c, and (b) to exploit complementary information
in
in the combined M/EEG and fMRI datasets
that refer to the same underlying brain activity subspace
c, and (b) to exploit complementary information
in  and
and  to gain new knowledge about c
and its underlying dynamics.
to gain new knowledge about c
and its underlying dynamics.
Methods for identifying the common subspace and exploiting
complementarity can utilize overlapping spatial, temporal,
or spatiotemporal information. Given that M/EEG
excels in the temporal domain and fMRI excels in the spatial
domain, we expect that an optimal approach should incorporate
both spatial and temporal information. All of the
spatial information in M/EEG comes from the locations
and geometry of the sensors, which we can co-register
with structural MRI. Methods for solving the M/EEG
inverse problem have this anatomical information at their
disposal, and can map source estimates to an anatomical
space in common with fMRI. Some studies have reported
convergent spatial results of independent M/EEG and fMRI
analyses (e.g., Grimm et al., 1998; Ball et
al., 1999). Other studies have sought to utilize
fMRI statistic images to constrain M/EEG inverse solutions
by dipole seeding (Ahlfors et al., 1999; Opitz
et al., 1999; Scherg et al., 1999) or regularization
of a linear estimator (Liu et al., 1998; Babiloni
et al., 1999). Wagner et al. (1999)
conducted a simulation study of both approaches.
An important goal of these methods is to use fMRI to bolster
confidence in the spatial aspect of M/EEG solutions so
that the latter can be used to extract source time series
on a millisecond time scale, which in turn can serve as
the basis for dynamic analysis of large-scale neural networks
(Thatcher et al., 1994). Ioannides (1999)
cautioned that statistic images obtained from fMRI (or
PET) block designs might be unsuitable for constraining
the analysis of MEG transients due to disparate temporal
scales. This criticism does not necessarily apply
to event-related fMRI designs. However, in relation
to the method presented in this paper, we note that (to
our knowledge) no approach that uses fMRI statistic images
to improve M/EEG source estimates has yet made explicit
use of the temporal information contained in the BOLD
signal: Although the datasets have been co-registered
in space, they have not been co-registered in time, i.e.,
temporally aligned. RS Menon et al. (1998)
have shown that relative BOLD signal onsets in different
regions may resolve timing sequences within tens of milliseconds.
Similarly, Wildgruber et al. (1997) associated
two phases of movement-related potentials (BP and motor
potential) with sequential event-related average hemodynamic
response onsets (in SMA and M1, respectively). V
Menon et al. (1997) used same-subject, same-task
event-related EEG and fMRI responses to gain information
about generators of the P300 ERP component. This
study demonstrated that temporal alignment of the linear
event-related EEG and fMRI responses is both possible
and useful. Guy et al. (1999) temporally
aligned EEG and fMRI responses with respect to periodic
visual stimuli, and found quasi-harmonic components of
the stimulus driving frequency in both modalities, which
demonstrate nonlinearities in the brain response.
The temporal alignment problem would vanish if fMRI acquisition
were truly simultaneous with M/EEG. Of course, this
is physically impossible with MEG. Ives et al.
(1993) demonstrated this possibility with ongoing EEG,
and Bonmasser et al. (1999) measured visual evoked
potentials concurrently with fMRI. Suitable controls
can effectively eliminate image artifacts caused by the
EEG apparatus (Krakow et al., 2000). Artifacts
in the EEG signal occur in the MR environment due to the
following sources: the static magnetic field interacts
with the EEG instrument; subject movements in the static
field induce currents; the ballistocardiogram (pulse)
artifact; mechanical vibrations; and especially inductive
effects of gradient switching during MR sequences (Lemieux
et al., 1999). Various approaches, some hardware
and others software, can reportedly deal with these issues.
Another unresolved issue is the possibility that the MR
environment may alter somewhat the EEG signal itself at
a physiological level. With reference to the right
half of Figure 1, we note that the cross-modal analyses
described here are pertinent to actual simultaneous datasets.
However, because we do not assume simultaneous M/EEG
and fMRI acquisitions, it is generally necessary to perform
temporal alignment. This, in turn, relies on inter-session
reliability of the task-related brain responses, which
is a testable assumption (Noll et al., 1997; Mangun
et al., 1998; Aguirre et al., 1998). The
remaining temporal alignment difficulties would vanish
if brain responses were strictly linear functions of event-related
task variables. Then same-type events would naturally
align their respective event-related averages (Menon et
al., 1997), which approximate linear impulse response
functions. However, this direct alignment approach
cannot handle dynamic nonlinearities, which are theoretically
important (Poggio and Reichardt, 1981; Friston, 2000)
and empirically observed (Vazquez and Noll, 1998; Friston
et al., 1998; Guy et al., 1999; Ances et
al., 2000). Furthermore, our strategy for cross-modal
analysis requires that we handle continuous, as well as
event-related, task variables. A viable tactic for
solving both the temporal alignment problem and the problem
of continuous task variables is to use nonlinear system
identification techniques (Schetzen, 1980; Bendat, 1998)
(a) to model M/EEG and fMRI responses and interactions
separately with respect to the same class of task realizations
(which may be either event-related or continuous) and
(b) to conduct virtual simultaneous experiments by driving
both nonlinear models in parallel with the same task realizations.
The approach detailed below is particularly indebted
to Friston et al. (1998), who (a) demonstrated
the viability of the pure Volterra approach for modeling
fMRI time series using second order kernels, (b) conducted
a virtual experiment with the model, and (c) placed the
Volterra model in the familiar context of the general
linear model. As an example of (c), the authors
constructed SPM{F} images of voxels with significant nonlinear
interactions. Furthermore, the Volterra approach
naturally extends to the analysis of effective connectivity
and modulatory interactions in large-scale neural networks
(Friston and Büchel, 2000) by replacing the task variables
with inputs from other brain regions.
2. Identification of Volterra-Based
System Models
In this section, we describe methods
for identifying the parameters of the three nonlinear,
time-invariant system models represented by equations
(3), (4), and (5). We start with the pure Volterra
model (equation (6)), and subsequently extend to the differential-Volterra
model (equation (22)) and the self-history Volterra model
(equation (25)). These identification methods utilize
“raw” (not averaged or statistically processed)
continuous signals. To simplify the description
below, we assume that there are no temporal gaps in the
data. However, in practice, the data may consist
of multiple runs, and each run may contain deleted segments
due to artifact rejection. Suitable bookkeeping
of the time variable can handle these discontinuities.
3. The Pure Volterra Model
| |
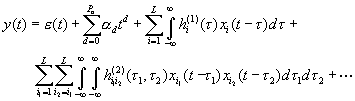 |
(6) |
Here  arises from a random background process,
which is primarily task-unrelated brain activity.
We have included low-order powers of time with coefficients
arises from a random background process,
which is primarily task-unrelated brain activity.
We have included low-order powers of time with coefficients
 in order to account for
in order to account for  slow drifts plus a constant, which
we regard as uninteresting. The experimentally driven
responses and interactions of interest are modeled by
the Volterra kernels
slow drifts plus a constant, which
we regard as uninteresting. The experimentally driven
responses and interactions of interest are modeled by
the Volterra kernels  (order 1) and
(order 1) and  (order 2). Task variables linearly
convolve with their first order kernels, and product pairs
of task variables bilinearly convolve with their second
order kernels. Higher order Volterra kernels n-linearly
convolve with product n-tuples of task variables;
however, the number of unknown parameters that we must
estimate increases sharply with the order (see equation
(12) ).
(order 2). Task variables linearly
convolve with their first order kernels, and product pairs
of task variables bilinearly convolve with their second
order kernels. Higher order Volterra kernels n-linearly
convolve with product n-tuples of task variables;
however, the number of unknown parameters that we must
estimate increases sharply with the order (see equation
(12) ).
A first order kernel  represents the linear response associated
with task variable
represents the linear response associated
with task variable  at time lag
at time lag  . We assume that responses are transient,
i.e., the kernel is zero except for a finite interval
of time lags. If the task variable corresponds with
a stimulus, then we assume that the brain response follows
the stimulus, which implies that a sensory kernel is identically
zero for all non-positive lags. If the task variable
corresponds with a behavior, then we assume that the brain
“response” precedes the behavior, which
implies that a motor kernel is identically zero for all
non-negative lags. For tasks that involve tight
coupling between a stimulus and a behavior, we can allow
sensorimotor kernels that are nonzero for both positive
and negative lags. Whenever possible, however, we
prefer to preserve causality by teasing apart the sensory
and motor responses. In general, we assume that
each task variable has an associated effective range
of time lags, such that a kernel is identically zero outside
the effective range.
. We assume that responses are transient,
i.e., the kernel is zero except for a finite interval
of time lags. If the task variable corresponds with
a stimulus, then we assume that the brain response follows
the stimulus, which implies that a sensory kernel is identically
zero for all non-positive lags. If the task variable
corresponds with a behavior, then we assume that the brain
“response” precedes the behavior, which
implies that a motor kernel is identically zero for all
non-negative lags. For tasks that involve tight
coupling between a stimulus and a behavior, we can allow
sensorimotor kernels that are nonzero for both positive
and negative lags. Whenever possible, however, we
prefer to preserve causality by teasing apart the sensory
and motor responses. In general, we assume that
each task variable has an associated effective range
of time lags, such that a kernel is identically zero outside
the effective range.
A second order kernel  represents the bilinear interaction
between task variables
represents the bilinear interaction
between task variables  and
and  at respective time lags
at respective time lags  and
and  . Kernels with
. Kernels with  represent quadratic self-interactions.
We assume that a second order kernel can be nonzero only
when both time lags are within the effective ranges of
their associated task variables.
represent quadratic self-interactions.
We assume that a second order kernel can be nonzero only
when both time lags are within the effective ranges of
their associated task variables.
| |
 |
(7) |
| |
 |
(8) |
| |
 |
(9) |
| |
 |
(10) |
| |
 |
(11) |
Because we obtain the  functions by convolving known task
variables with predetermined basis functions, we thus
can jointly estimate, via generalized least squares, the
a
(drift), b
(linear response), and g (nonlinear interaction) parameters.
Equations (7) and (8) reconstruct the first and second
order Volterra kernels from these parameters. We
note that the self-interaction kernels are symmetric.
functions by convolving known task
variables with predetermined basis functions, we thus
can jointly estimate, via generalized least squares, the
a
(drift), b
(linear response), and g (nonlinear interaction) parameters.
Equations (7) and (8) reconstruct the first and second
order Volterra kernels from these parameters. We
note that the self-interaction kernels are symmetric.
4. Gauss-Markov Estimation of Volterra Kernel Parameters
The total number  of estimated parameters for the second
order Volterra model is
of estimated parameters for the second
order Volterra model is
| |
 |
(12) |
This total includes a constant,
the a
trend coefficients, the b response parameters, the
symmetric self-interaction g parameters, and the cross-interaction
g parameters, from which we
form a vector  of length
of length 
| |
 |
(13) |
Corresponding with the column
vector  , we form a row vector
, we form a row vector  of length
of length  that reflects the experimental situation
at time
that reflects the experimental situation
at time  (in terms of basis-convolved task variables):
(in terms of basis-convolved task variables):
| |
 |
(14) |
For an experimental run consisting
of  time samples, the full
time samples, the full  design matrix
design matrix  is
is
| |
 |
(15) |
Collecting the measured signals
for all time samples yields a data vector  of length
of length  :
:
| |
 |
(16) |
Likewise, we form an error vector
 of length
of length  :
:
| |
 |
(17) |
We may rewrite equation (11)
in matrix form as
| |
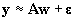 |
(18) |
The time series  has a serial correlation structure
that derives from “background” neural activity,
other physiological activity, motion effects, and/or scanner/system
noise (Zarahn et al., 1997). This, in turn,
implies that the frequency spectrum is not flat (i.e.,
has a serial correlation structure
that derives from “background” neural activity,
other physiological activity, motion effects, and/or scanner/system
noise (Zarahn et al., 1997). This, in turn,
implies that the frequency spectrum is not flat (i.e.,
 is not white noise). If the error
process is approximately gaussian and stationary, we can
solve (18) via ordinary least squares estimation if we
first prewhiten the data. In the frequency domain,
we divide by the amplitude at each frequency, which flattens
(“whitens”) the spectrum. The time domain
equivalent of prewhitening can be formally written as
equation (19), where
is not white noise). If the error
process is approximately gaussian and stationary, we can
solve (18) via ordinary least squares estimation if we
first prewhiten the data. In the frequency domain,
we divide by the amplitude at each frequency, which flattens
(“whitens”) the spectrum. The time domain
equivalent of prewhitening can be formally written as
equation (19), where is the
is the  variance-covariance matrix for the
error time series. Stationarity of the background
activity implies that each autocovariance
variance-covariance matrix for the
error time series. Stationarity of the background
activity implies that each autocovariance  depends only on the lag
depends only on the lag  . If we extrapolate autocovariances
beyond a specified lag
. If we extrapolate autocovariances
beyond a specified lag  using the principle of maximum entropy,
then we can use autoregressive (AR) methods to design
a prewhitening filter in the time domain by inverting
an AR model of the error process, which involves only
autocovariances with lags less than
using the principle of maximum entropy,
then we can use autoregressive (AR) methods to design
a prewhitening filter in the time domain by inverting
an AR model of the error process, which involves only
autocovariances with lags less than  (Friston, Josephs et al., 2000;
Appendix A). Applying the prewhitening filter
(Friston, Josephs et al., 2000;
Appendix A). Applying the prewhitening filter  to each term of equation (18), we obtain
to each term of equation (18), we obtain
| |
 |
(19) |
Under the assumptions,  is identically and independently distributed,
in which case the Gauss-Markov estimate for the parameters
is
is identically and independently distributed,
in which case the Gauss-Markov estimate for the parameters
is
| |
 |
(20) |
Here the “+” superscript
represents the pseudoinverse operation.
In practice, we typically must
use the same data to estimate the autocovariances of the
background brain activity with lags less than  . A feasible generalized least squares
approach (Dale et al., 2000) obtains a series of
estimates,
. A feasible generalized least squares
approach (Dale et al., 2000) obtains a series of
estimates,  , where
, where  is computed using
is computed using  (or another initial estimate);
(or another initial estimate); 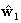 is computed using the approximation
is computed using the approximation
 , where we form
, where we form 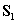 by calculating lag covariances from
the residual vector
by calculating lag covariances from
the residual vector  ; and so on. This series converges
to mutually consistent estimates of
; and so on. This series converges
to mutually consistent estimates of  and
and  , given the data and the modeling assumptions.
Friston, Josephs et al. (2000) have recently discussed
caveats regarding the effect of prewhitening filters on
statistical bias, which we discuss further below.
, given the data and the modeling assumptions.
Friston, Josephs et al. (2000) have recently discussed
caveats regarding the effect of prewhitening filters on
statistical bias, which we discuss further below.
Although not the only viable
approach for solving equation (20), singular value decomposition
(SVD) of  conveniently provides (a) a solution
that is stable, and (b) standard errors in the fitted
parameters, which reflect goodness-of-fit of the model
to the data (Press et al., 1992, §15.4).
Clearly, this analysis fits the general linear model (GLM).
As Friston et al. (1998) have demonstrated, the
statistical machinery of the GLM can be combined with
Volterra models to construct SPM{F} maps (Worsley 1994)
of (for example) brain regions that have significant nonlinear
kernels.
conveniently provides (a) a solution
that is stable, and (b) standard errors in the fitted
parameters, which reflect goodness-of-fit of the model
to the data (Press et al., 1992, §15.4).
Clearly, this analysis fits the general linear model (GLM).
As Friston et al. (1998) have demonstrated, the
statistical machinery of the GLM can be combined with
Volterra models to construct SPM{F} maps (Worsley 1994)
of (for example) brain regions that have significant nonlinear
kernels.
4.1 Tailoring of Basis Functions
The total number of estimated
parameters  of equation (12) depends primarily
on the number of task variables
of equation (12) depends primarily
on the number of task variables  and the number of basis functions utilized
per task variable. We wish to minimize
and the number of basis functions utilized
per task variable. We wish to minimize  in order to obtain stable estimates
from a limited number of data samples. However,
in opposition to this goal, we also seek to minimize the
approximation error of equations (7) and (8). For
fixed
in order to obtain stable estimates
from a limited number of data samples. However,
in opposition to this goal, we also seek to minimize the
approximation error of equations (7) and (8). For
fixed  (an index of task complexity), decreasing
the number of basis functions decreases
(an index of task complexity), decreasing
the number of basis functions decreases  , whereas increasing the number of basis
functions decreases approximation error. To optimize
this tradeoff, we strive to maximize the ability of a
given number of basis functions to represent Volterra
kernels. Since each set of basis functions is associated
with a task variable and its first order kernel, we take
an empirical approach that tailors the representation
of the first order kernel. We base this approach
on the observation that an average event-related response
approximates the impulse response (which is the first
order kernel) so long as the task events occur in temporal
isolation. Thus, if we have “calibration data”
for each task variable, consisting of isolated task events,
then we can make an event-related average to which we
tailor a set of parameterized basis functions. In
particular, we propose to use
, whereas increasing the number of basis
functions decreases approximation error. To optimize
this tradeoff, we strive to maximize the ability of a
given number of basis functions to represent Volterra
kernels. Since each set of basis functions is associated
with a task variable and its first order kernel, we take
an empirical approach that tailors the representation
of the first order kernel. We base this approach
on the observation that an average event-related response
approximates the impulse response (which is the first
order kernel) so long as the task events occur in temporal
isolation. Thus, if we have “calibration data”
for each task variable, consisting of isolated task events,
then we can make an event-related average to which we
tailor a set of parameterized basis functions. In
particular, we propose to use  basis functions that have the form
of a Poisson distribution:
basis functions that have the form
of a Poisson distribution:
| |
 |
(21) |
where  ;
;  is a “characteristic latency”
that roughly corresponds to the peak latency of the event-related
average response; and
is a “characteristic latency”
that roughly corresponds to the peak latency of the event-related
average response; and  is a “characteristic delay”
that roughly corresponds with the onset of the response.
The fundamental basis function (
is a “characteristic delay”
that roughly corresponds with the onset of the response.
The fundamental basis function ( ) can fit a “main peak”; the
) can fit a “main peak”; the
 functions can fit early oscillations
that occur before
functions can fit early oscillations
that occur before  ; and the
; and the  functions can fit slow waves that occur
after
functions can fit slow waves that occur
after  . For each number
. For each number  there are optimal values of
there are optimal values of  and
and  for approximating the average response.
Thus, for each task variable, we seek the smallest
for approximating the average response.
Thus, for each task variable, we seek the smallest  that produces an adequate approximation.
that produces an adequate approximation.
4.2 The Differential-Volterra Model
The differential-Volterra model extends the Volterra
model (equation (6)) by applying a differential operator
 to the data term
to the data term on the left-hand side (LHS) of equation
(6):
on the left-hand side (LHS) of equation
(6):
| |
 |
(22) |
where
| |
 |
(23) |
That is,
| |
 |
(24) |
Here  is the highest derivative (order) of
the differential operator
is the highest derivative (order) of
the differential operator  , which has constant coefficients
, which has constant coefficients  . The
. The  parameters are the first-degree (linear)
coefficients; the
parameters are the first-degree (linear)
coefficients; the  parameters are the second-degree (quadratic-bilinear)
coefficients; etc. The revised LHS is a derived
data term, obtained by forming linear and higher degree
combinations of the measured time series with its own
temporal derivatives. To prevent the derived data
term from vanishing, we require
parameters are the second-degree (quadratic-bilinear)
coefficients; etc. The revised LHS is a derived
data term, obtained by forming linear and higher degree
combinations of the measured time series with its own
temporal derivatives. To prevent the derived data
term from vanishing, we require  while considering order
while considering order  . Thus, the pure Volterra model of
equation (6) is the special case of equation (22) when
the degree is 1 and the order
. Thus, the pure Volterra model of
equation (6) is the special case of equation (22) when
the degree is 1 and the order  is 0 (noting that
is 0 (noting that  ). Whereas the error term
). Whereas the error term  for equation (6) is the error of the
original measured data, we note that the error term for
equation (22) is the error of the derived data.
For a first-degree operator, there are
for equation (6) is the error of the
original measured data, we note that the error term for
equation (22) is the error of the derived data.
For a first-degree operator, there are  undetermined linear coefficients.
For a second-degree operator, there are, in addition,
undetermined linear coefficients.
For a second-degree operator, there are, in addition,
 quadratic coefficients, and
quadratic coefficients, and  bilinear coefficients. If the
order
bilinear coefficients. If the
order  , for example, then the number of unknown
, for example, then the number of unknown
 parameters is 2 for a first-degree
operator, and 8 for a second-degree operator.
parameters is 2 for a first-degree
operator, and 8 for a second-degree operator.
A differential operator applied
to the data permits us, using relatively few additional
parameters, to complement the model of the driving
process (which is given on the RHS) by specifying
a model of the driven process (which is given on
the LHS). In short, the RHS specifies how to drive
some process, the nature of which the LHS
specifies. For example, the pure Volterra model
of equation (6) assumes that the brain process driven
by the task variables is identical to our measurement
process. However, it is equally plausible to conjecture
that the task variables drive changes in the brain,
as reflected by first derivatives of the measurement process.
If the original measurements and their derivatives are
both plausible candidates for the driven brain process,
then any linear combination of the two is yet another
plausible candidate. We can extend this to higher
order derivatives. Higher degree combinations give
the differential operator a nonlinear character, which
(as we noted above) may have theoretical significance
by permitting the deterministic and stochastic terms on
the RHS to interact. These considerations motivate the
rationale for using the differential operator of equation
(23) to specify the nature of the driven process.
Although Gauss-Markov methods
cannot directly estimate the l parameters, i.e., we cannot
simply append them to the  vector of equation (13), we can use
the identical methods detailed above as part of an iterative
approach. The steps of this procedure are as follows
(for a given degree and order
vector of equation (13), we can use
the identical methods detailed above as part of an iterative
approach. The steps of this procedure are as follows
(for a given degree and order  , starting with degree 1 and order 0):
, starting with degree 1 and order 0):
1.
Numerically evaluate the time derivatives  from the data
from the data  . Initially set
. Initially set  , and set all other coefficients to zero.
, and set all other coefficients to zero.
2.
Use the Gauss-Markov methods detailed above, substituting
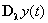 for
for  , to obtain an estimate
, to obtain an estimate  for the RHS parameters.
for the RHS parameters.
3.
Compute the modeled time series  , which we obtain by applying the design
matrix to the RHS parameter estimate and evaluating at
each time
, which we obtain by applying the design
matrix to the RHS parameter estimate and evaluating at
each time  .
.
4.
Use numerical methods to solve the ordinary differential
equation  for
for  .
.
5.
Compute the mean square error of the residual,
 .
.
6.
Employ a suitable, general minimization method
that iterates steps 2-5 to find  with minimum
with minimum  , subject to
, subject to  . We note that with low degree and
order,
. We note that with low degree and
order,  consists of relatively few parameters
compared with the number
consists of relatively few parameters
compared with the number  of Volterra kernel parameters.
of Volterra kernel parameters.
By design, the above steps start
with the basic Volterra model (equation (6)), and generally
will find a differential-Volterra model (equation (22))
with smaller residual error. For low degree and
order, we can attribute any improvement gained to the
use of just a few additional parameters (relative to the
number  of Volterra kernel parameters), which
effectively change what the Volterra kernels are modeling.
Thus with moderation, we expect that this procedure should
not risk overmodeling the data. This point is important
in regard to the objective of finding improved models
of the data that generalize across acquisition
sessions, i.e., we must avoid models that are overly specific
to one session. Thus, until we have characterized
these methods, it will be necessary to acquire multiple
session datasets for each modality in order to determine
model order-degree that optimizes generalization across
sessions.
of Volterra kernel parameters), which
effectively change what the Volterra kernels are modeling.
Thus with moderation, we expect that this procedure should
not risk overmodeling the data. This point is important
in regard to the objective of finding improved models
of the data that generalize across acquisition
sessions, i.e., we must avoid models that are overly specific
to one session. Thus, until we have characterized
these methods, it will be necessary to acquire multiple
session datasets for each modality in order to determine
model order-degree that optimizes generalization across
sessions.
Steps 4 and 6 depend on unspecified
numerical methods for solving ordinary differential equations
and minimization of multidimensional functions, respectively.
Here we are contemplating a small number of differential
parameters (1-8), so we do not expect that the choice
of minimization method is critical. We can use analytic
methods to solve second order linear (first-degree) differential
equations (two unknown coefficients), and Laplace transform
methods for higher order linear equations. The quadratic-bilinear
(second-degree) equations call for different methods,
which we do not discuss here.
4.3 The Self-History Volterra Model
The self-history Volterra model extends the pure Volterra
model (equation (6)) by including the past of the response
variable  in the Volterra expansion for the present:
in the Volterra expansion for the present:
| |
 |
(25) |
Here we have added a linear self-response kernel, a quadratic
self-interaction kernel, and bilinear task-response interaction
kernels. Unlike the differential-Volterra model
of equation (22), we can append the new parameters of
the self-history Volterra model to the  vector of equation (13). Although
we cannot empirically “calibrate” basis functions
as described above, the Laguerre functions are a natural
basis for expanding these new kernels (Schetzen, 1980).
One caveat of the self-history approach is the introduction
of measurement errors into the input variables (Press
et al., 1992, §15.3). Equation (12) gives
the number of parameters to estimate, with the understanding
that there is the equivalent of one extra task variable.
vector of equation (13). Although
we cannot empirically “calibrate” basis functions
as described above, the Laguerre functions are a natural
basis for expanding these new kernels (Schetzen, 1980).
One caveat of the self-history approach is the introduction
of measurement errors into the input variables (Press
et al., 1992, §15.3). Equation (12) gives
the number of parameters to estimate, with the understanding
that there is the equivalent of one extra task variable.
Self-history Volterra model identification is computationally
simpler than differential-Volterra model identification,
although the latter typically requires fewer extra parameters
than the former. We can combine both models to make
a self-history differential-Volterra model, which has
the form of equation (1). In practice, the key consideration
for choosing from among the four model types (pure Volterra,
differential-Volterra, self-history Volterra, and self-history
differential-Volterra) is the ability to efficiently generalize
across sessions, i.e., the ability to obtain a good fit
of data acquired in one session using model parameters
identified with a different session’s data.
All else being equal, we prefer the simplest model.
(We do not address these empirical issues here.)
Task-Related and Cross-Modal Analyses
At this point, we can implement the strategy summarized
in Figure 1, where we substitute one of the four model
types for “system model”. If the task-related
fMRI and M/EEG models (on the left side of Figure 1) efficiently
generalize across sessions, then we can drive them simultaneously
with the same simulated task variables to generate a virtual
simultaneous dataset. Although we used the stochastic
component to identify the system models, we can omit this
during the simulation. In this case, the simulated
data will reflect only the deterministic response, which
is free from the influences of “brain noise”.
The goal of fMRI-related M/EEG analysis at a particular
location is to determine a set of channel weights such
that the derived M/EEG signal has maximum dynamic coupling
with the fMRI BOLD signal. We accomplish this in
the context of identifying a system model, where the input
is a BOLD signal at the location (perhaps averaged in
a local neighborhood). The measured signal  in equations (6), (22), or (25) is replaced
by
in equations (6), (22), or (25) is replaced
by
| |
 |
(26) |
where  is a M/EEG channel vector at time
is a M/EEG channel vector at time  , and
, and  is a row of channel weights that we
aim to determine. More generally,
is a row of channel weights that we
aim to determine. More generally,  may contain up to three rows that permit
up to three source current orientations at the location.
The norm operation converts the resultant one-to-three
dimensional vector to a nonnegative number, which implicitly
assumes that the BOLD signal is not orientation sensitive.
may contain up to three rows that permit
up to three source current orientations at the location.
The norm operation converts the resultant one-to-three
dimensional vector to a nonnegative number, which implicitly
assumes that the BOLD signal is not orientation sensitive.
We can estimate  iteratively, as follows. On the
first iteration, we set
iteratively, as follows. On the
first iteration, we set  to a theoretical estimator of local
activity, such as a beamformer (e.g., Sekihara et al.,
2000). Having fixed
to a theoretical estimator of local
activity, such as a beamformer (e.g., Sekihara et al.,
2000). Having fixed  , we calculate
, we calculate  , identify a system model with respect to
the BOLD signal input, and measure goodness-of-fit.
On subsequent iterations, a multidimensional minimization
method searches for
, identify a system model with respect to
the BOLD signal input, and measure goodness-of-fit.
On subsequent iterations, a multidimensional minimization
method searches for  parameters that optimize goodness-of-fit.
The associated best-fitting model maximizes the dynamic
coupling between the BOLD signal and the source activity
estimated by (26). In practice, we can substantially
reduce the number of unknown parameters, i.e., the elements
of
parameters that optimize goodness-of-fit.
The associated best-fitting model maximizes the dynamic
coupling between the BOLD signal and the source activity
estimated by (26). In practice, we can substantially
reduce the number of unknown parameters, i.e., the elements
of  , by representing
, by representing  as the sum of a few principal components.
as the sum of a few principal components.
The goal of M/EEG-related fMRI analysis is to provide
a more precise temporal context for statistical detection
of activations in space by modeling the local BOLD response
system using input obtained from the M/EEG-derived signal
of equation (26). The local BOLD signal depends
on three main physiological variables: local oxygen metabolic
rate, which is a consequence of neuronal activity; regional
cerebral blood flow, which is regulated by neuronal activity;
and regional cerebral blood volume, which depends on the
blood flow (Buxton, 1998). Thus, the local hemodynamic/metabolic
system receives neuronal activity as input and produces
BOLD signal as output. “Activation”
means that this system is active, i.e., that there is
significant dynamic coupling between the input neuronal
activity and the output BOLD signal. We can test
the hypothesis of significant activation in the context
of dynamic modeling by forming an F-statistic as the ratio
of (a) the magnitude of the projection of the data into
the model space over (b) the magnitude of the residual
error (Worsley and Friston, 1995; Buxton et al.,
2000). However, we cannot directly model the hemodynamic/metabolic
system with respect to the unobserved neuronal activity.
Because the known task variables drive the unknown neuronal
activities of interest, we typically must replace the
latter with the former as input to the dynamic model.
On the other hand, these sensory and motor task variables
are at least one stage removed from the actual neuronal
activity of a brain region. More typically, the
neuronal activities of other brain regions intervene.
Moreover, whereas the task variables are uniform across
voxels, actual neuronal activity varies from region to
region. Therefore, it may be highly beneficial to
replace the task variables with region-by-region estimates
of neuronal activity, such as those derived from M/EEG.
Although imperfect, these temporally improved local estimates
of the task-related neuronal activity may yield increased
statistical power for detecting activated voxels, especially
for brain regions that are most removed from the sensory
and motor variables.
5. Discussion
In brief, we have outlined a solution method for problem
A that uses the framework of nonlinear system identification
to simulate virtual simultaneous data, and then to conduct
cross-modal analyses. The fMRI-related M/EEG analysis
permits us to “tune” an inverse estimator
for source activity at a location. Although BOLD
responses lag behind the neuronal responses that M/EEG
detects, we nevertheless can formally model the M/EEG
signal as an output to a dynamic system with a BOLD signal
input. This is not a causal model, but the machinery
permits us to empirically derive an estimator of source
activity that maximizes the local dynamic coupling between
BOLD and M/EEG signals. M/EEG-related fMRI analysis
models the BOLD signal as the output of a local hemodynamic/metabolic
system that has M/EEG-derived signals as inputs.
This system is presumably causal, and it permits us to
estimate the possibly nonlinear hemodynamic responsiveness
of fMRI to neuronal activity.
We have not here directly addressed problem B, which
is to use both sets of measurements to jointly estimate
the underlying dynamics, which, in turn, requires a joint
spatiotemporal estimate of the neuronal activity.
However, we have made some progress toward problem B by
virtue of attending to problem A. In particular,
the state dynamics equation (1) has motivated our approach,
and we have made some theoretical progress toward characterizing
the observation equations (2): The cross-modal analysis
produces an fMRI-restricted version of the inverse M/EEG
transfer function  (embodied in the matrix
(embodied in the matrix  ) and an M/EEG-restricted version of the
forward fMRI BOLD transfer function
) and an M/EEG-restricted version of the
forward fMRI BOLD transfer function  (embodied in the nonlinear system model
of the M/EEG-related fMRI analysis). These transfer
functions are mutually restricted in the sense that they
are relative to each other, having both been “tuned”
to the common subspace
(embodied in the nonlinear system model
of the M/EEG-related fMRI analysis). These transfer
functions are mutually restricted in the sense that they
are relative to each other, having both been “tuned”
to the common subspace  of neuronal activity that is strongly
coupled with the measured signals of both modalities.
Nevertheless, we are positioned to contemplate a “simultaneous
solution” to equations (1) and (2).
of neuronal activity that is strongly
coupled with the measured signals of both modalities.
Nevertheless, we are positioned to contemplate a “simultaneous
solution” to equations (1) and (2).
The question of optimal experimental design for model
identification (parameter estimation) and associated statistics
(signal detection) has been a recent topic of discussion
in the fMRI literature. Efficient parameter estimation
utilizes randomized, short-latency inter-event intervals
(Dale, 1999); the conventional block design with fixed
intervals is optimal for statistical signal detection
(Friston et al., 1999); and a hybrid block-probability
approach represents a compromise (Friston et al.,
1999; Buxton et al., 2000). Discussions of
the estimation-detection tradeoff have considered discrete
designs that consist of binary point processes of events.
Identification of continuous-input physiological systems
via white noise analysis (e.g., Marmarelis and Naka, 1974)
is a purely random approach, which parallels the efficient
estimators for discrete, binary inputs. Experimental
design via white noise inputs clearly represents the “least
designed design” that attempts to sample the entire
input space without bias. However, actual neurons
and organisms have extremely biased interests in “remote
corners” of input space (Lettvin et al.,
1959; Hubel and Wiesel, 1959), and tend to “tune
out” random processes. Because statistical
signal detection relies on robust responses, these considerations
suggest that we ultimately may desire an adaptive, real-time
approach to task design that combines random “importance
sampling” with functional hypothesis testing (problem
C). That is, in the context of an explicit hypothesis
about an objective function that a brain region putatively
optimizes, we might use online statistical signal detection
to identify a subspace of task variables that we infer
are interesting to a brain region after observing its
robust response. Such an adaptive importance sampling
design should dynamically resolve the discrepancy between
optimal estimation and optimal detection (which discrepancy
signals that the current design is not functionally
optimal).
A parallel issue concerns the effect of using (or not
using) estimated serial correlations of the background
noise on (i) the efficiency of parameter estimation versus
(ii) robustness of signal detection. The Gauss-Markov
estimator described above makes use of estimated temporal
covariances for more efficient parameter estimation; however,
the associated signal detection statistics are not robust
with respect to errors in the temporal covariance estimates
(Friston, Josephs et al., 2000). A key reason
for this estimation-detection conflict is the requirement
of gaussian field theory to use the same statistical model
across all voxels (Worsley, 1994; Worsley and Friston,
1995), whereas temporal covariances of the background
activity are spatially nonstationary (Zarahn et al., 1997).
However, except for the final step of forming a statistic
image, our primary concern here is with a third criterion,
which we can evaluate empirically: (iii) within-subject,
between-session test-retest reliability of the dynamic
models (Genovese et al., 1997; Noll et al.,
1997). Within-session efficiency of parameter estimation
does not guarantee that a model will generalize across
sessions. Only inter-session stability can warrant
the use of specific dynamic modeling features (such as
voxel-specific noise estimates, differential operators,
or self-history kernels) for conducting virtual simultaneous
experiments and cross-modal analyses.
References
Ahlfors SP, Simpson GV, Dale AM, Belliveau
JW, Liu AK, Korvenoja A, Virtanen J, Huotilainen M, Tootell
RB, Aronen HJ, Ilmoniemi RJ (1999): Spatiotemporal activity
of a cortical network for processing visual motion revealed
by MEG and fMRI. J Neurophysiol 82(5):
2545-55.
Aguirre GK, Zarahn E, D’Esposito M (1998):
The variability of human, BOLD hemodynamic responses.
Neuroimage 8: 360-9.
Ances BM, Zarahn E, Greenberg JH, Detre JA
(2000): Coupling of neural activation to blood flow in
the somatosensory cortex of rats is time-intensity separable,
but not linear. J Cereb Blood Flow Metab 20(6): 921-30.
Arbib MA, Érdi
P, Szentágothai J (1998): Neuronal Organization: Structure,
Function, and Dynamics. Cambridge, MA:
The MIT Press.
Arthur RM, Geselowitz DB (1970): Effects of
inhomogeneities on the apparent location and magnitude
of a cardiac current dipole generator. IEEE
Trans Biomed Eng 17:141-6.
Ashby WR (1960):
Design for a Brain. London: Science
Paperbacks.
Awada, KA and Greenblatt, RE. (1999): EEG source
localization conductivity sensitivity using a 3D finite
element model. Neuroimage 9:S138.
Babiloni F, Carducci F, Del Gratta C, Roberti
GM, Cincotti F, Bagni O, Romani GL, Rossini PM, Babiloni
C (1999): Multimodal integration of high resolution EEG,
MEG and functional magnetic resonance data. Int
J Bioelectromagnetism 1(1).
Ball T, Schreiber A, Feige B, Wagner M, Lucking
CH, Kristeva-Feige R (1999): The role of higher-order
motor areas in voluntary movement as revealed by high-resolution
EEG and fMRI. Neuroimage 10: 682-94.
Bendat JS (1998):
Nonlinear System Techniques and Applications.
New York: John Wiley.
Bonmassar G, Anami K, Ives J, Belliveau JW
(1999): Visual evoked potential (VEP) measured by simultaneous
64-channel EEG and 3T fMRI. Neuroreport 10: 1893-7.
Brandt ME, Jansen BH (1991): The relationship
between prestimulus alpha amplitude and visual evoked
potential amplitude. Int J Neurosci 61: 261-68.
Buxton RB, Liu TT, Martinez A, Frank LR, Luh
W-M, Wong EC (2000): Sorting out event-related paradigms
in fMRI: the distinction between detecting an activation
and estimating the hemodynamic response. Neuroimage 11: S457.
Buxton RB, Wong EC, Frank LR (1998): Dynamics
of blood flow and oxygenation changes during brain activation:
the balloon model. Magn Reson Med 39: 855-64.
Dale AM (1999): Optimal experimental design
for event-related fMRI. Hum
Brain Mapp 8:
109-14.
Dale AM, Liu AK, Fischl BR, Buckner RL, Belliveau
JW, Lewine JD, Halgren E (2000): Dynamic statistical parametric
mapping: combining fMRI and MEG for high-resolution imaging
of cortical activity. Neuron 26(1): 55-67.
D’Esposito M, Zarahn E, Aguirre GK, Rypma
B (1999): The effect of normal aging on the coupling of
neural activity to the BOLD hemodynamic response.
Neuroimage 10: 6-14.
Friston KJ (1997): Another neuronal code?
Neuroimage 5: 213-20.
Friston KJ (2000): The labile brain.
I. Neuronal transients and nonlinear coupling. Phil
Trans R Soc Lond B 355: 215-236.
Friston KJ, Büchel C (2000): Attentional
modulation of effective connectivity from V2 to V5/MT
in humans. Proc Natl Acad Sci USA 97(13): 7591-7596.
Friston KJ, Josephs O, Rees G, & Turner
R (1998): Nonlinear event-related responses in fMRI.
Magn Reson Med 39:41-52.
Friston JK, Josephs O, Zarahn E, Holmes AP,
Rouquette S, Poline J (2000): To smooth or not to smooth?
Bias and efficiency in fMRI time-series analysis.
Neuroimage 12(2): 196-208.
Friston KJ, Zarahn E, Josephs O, Henson RN,
Dale AM (1999): Stochastic designs in event-related fMRI.
Neuroimage 10: 607-19.
Genovese CR, Noll DC, Eddy WF (1997): Estimating
test-retest reliability in functional MR imaging. I: Statistical
methodology. Magn Reson Med 38(3): 497-507.
Grimm C, Schreiber A, Kristeva-Feige R, Mergner
T, Hennig J, Lucking CH (1998): A comparison between electric
source localisation and fMRI during somatosensory stimulation.
Electroencephalogr Clin Neurophysiol 106: 22-9.
Guy CN, ffytche DH, Brovelli A, Chumillas J
(1999): fMRI and EEG responses to periodic visual stimulation.
Neuroimage 10: 125-48.
Hämäläinen M and Sarvas J (1987): Feasibility
of the homogeneous head model in the interpretation of
neuromagnetic fields. Physics
Med Biol 32:91-98.
Helmholtz HLF (1853): Ueber einige Gesetze
der Vertheilung elektrischer Ströme in körperlichen Leitern
mit Anwendung auf die thierisch-elektrischen Versuche.
Ann Physik und Chemie 89: 211-33, 354-77.
Hubel DH, Wiesel TN (1959): Receptive fields
of single neurones in the cat’s striate cortex.
J Physiol 148: 574-591.
Ioannides AA (1999): Problems associated with
the combination of MEG and fMRI data: theoretical basis
and results in practice. In: T Yoshimoto, M Kotani,
S Kuriki, H Karibe, N Nakasato (eds), Recent
Advances in Biomagnetism. Sendai: Tohoku
University Press, 133-6.
Isaka S, Sebald V (1993): Control strategies
for arterial blood pressure regulation. IEEE
Trans Biomed Eng 40: 353-363.
Ives JR, Warach S, Schmitt F, Edelman RR, Schomer
DL (1993): Monitoring the patient’s EEG during echo
planar MRI. Electroencephalogr Clin Neurophysiol 87: 417-20.
Kelso JAS (1995):
Dynamic Patterns: The Self-Organization of Brain and
Behavior. Cambridge, MA: MIT Press.
Klingelhofer J, Hajak G, Matzander G, Schulz-Varszegi
M, Sander D, Ruther E, Conrad B (1995): Dynamics of cerebral
blood flow velocities during normal human sleep.
Clin Neurol Neurosurg
97: 142-8.
Koch K (1999): Biophysics of Computation:
Information Processing in Single Neurons. New
York: Oxford University Press.
Krakow K, Allen PJ, Symms MR, Lemieux L, Josephs
O, Fish DR (2000): EEG recording during fMRI experiments:
image quality. Hum Brain Mapp10(1): 10-5.
Kwong KK, Belliveau JW, Chesler DA, Goldberg
IE, Weisskoff RM, Poncelet BP, Kennedy DN, Hoppel BE,
Cohen MS, Turner R, Cheng H-M, Brady TJ, Rosen BR (1992):
Dynamic magnetic resonance imaging of human brain activity
during primary sensory stimulation. Proc Natl Acad Sci USA 89: 5675-5679.
Lemieux L, Allen PJ, Krakow K, Symms MR, Fish
DR (1999): Methodological issues in EEG-correlated functional
MRI experiments. Int J Bioelectromagnetism 1(1).
Lettvin JY, Maturana HR, McCulloch WS, Pitts
WH (1959): What the frog’s eye tells the frog’s
brain. 11.0pt'>Proc IRE 47: 1940-1951.
Liu AK, Belliveau JW, Dale AM (1998): Spatiotemporal
imaging of human brain activity using functional MRI constrained
magnetoencephalography data: Monte Carlo simulations.
Proc Natl Acad Sci USA 95: 8945-50.
Lorente de Nó R (1947): A Study of Nerve Physiology.
New York: Rockefeller Institute for Medical Research.
Malmivuo J, Plonsey R (1995). Bioelectromagnetism. New York:
Oxford University Press.
Mangun GR, Buonocor MH, Girelli M, Jha AP (1998):
ERP and fMRI measures of visual spatial selective attention.
Hum Brain Mapp 6: 383-9.
Marmarelis PZ, Naka KI (1974): Identification
of multi-input biological systems. IEEE Trans Biomed Eng 21: 88-101.
Menon RS, Luknowsky DC, Gati JS (1998): Mental
chronometry using latency-resolved functional MRI.
Proc Natl Acad Sci USA 95: 10902-7.
Menon R, Ogawa S, Hu X, Strupp JP, Anderson
P, Ugurbil K (1995): BOLD based functional MRI at 4 Tesla
includes a capillary bed contribution: echo-planar imaging
correlates with previous optical imaging using intrinsic
signals. Magn
Reson Med 33: 453-9.
Menon V, Ford JM, Lim KO, Glover GH, Pfefferbaum
A (1997): Combined event-related fMRI and EEG evidence
for temporal-parietal cortex activation during target
detection. Neuroreport 8: 3029-37.
Noll DC, Genovese CR, Nystrom LE, Vazquez AL,
Forman SD, Eddy WF, Cohen JD (1997): Estimating test-retest
reliability in functional MR imaging. II: Application
to motor and cognitive activation studies. Magn
Reson Med 38(3): 508-17.
Nunez (1995):
Neocortical Dynamics and Human EEG Rhythms.
New York: Oxford University Press.
Ogawa S, Chen W, Zhu X-H, Ugurbil K (2000):
Probing neuronal events by fMRI at neuronal time scale.
Neuroimage 11: S562.
Ogawa S, Tank DW, Menon R, Ellermann JM, Kim
S-G, Merkle H, Ugurbil K (1992): Intrinsic signal changes
accompanying sensory stimulation: Functional brain mapping
with magnetic resonance imaging. Proc
Natl Acad Sci USA 89: 5951-5955.
Opitz B, Mecklinger A, von Cramon DY, Kruggel
F (1999): Combining electrophysiological and hemodynamic
measures of the auditory oddball. Psychophysiology
36(1): 142-7.
Ozaki, T (1985): Nonlinear time series models
and dynamical systems. In: Hannan EJ, Krishnaiah
PR and Rao MM (eds) Handb Statistics 5:25-83.
Palerm C, Kaufman H, Bequette W (1999): Validation
of an adaptive hemodynamic controller. Proceedings
of the First Joint BMES/EMBS Conference: 254.
Phillips WA, Singer W (1997): In search of common foundations
for cortical computation. Behavioral and Brain
Sciences 20: 657-722.
Pflieger ME (1991): A Theory of Optimal Event-Related
Brain Signal Processors Applied to Omitted Stimulus Data.
Doctoral dissertation, The Ohio State University.
Poggio T, Reichardt WE (1981): Characterization of nonlinear
interactions in the fly’s visual system. In:
WE Reichardt and T Poggio (eds), Theoretical Approaches
in Neurobiology. Cambridge, MA: The MIT Press,
64-84.
Pöppel E (1997): A hierarchical model of temporal perception.
Trends Cog Sci 1: 56-61.
Press WH, Teukolsky SA, Vetterling WT, Flannery BP (1992):
Numerical Recipes in C (2nd Edition).
Cambridge University Press.
Scherg M, Linden DEJ, Muckli L, Roth R, Drüen K, Ille
N, Linek S, Zanella FE, Singer W, Goebel R (1999): Combining
MEG with fMRI in studies of the human visual system.
In: T Yoshimoto, M Kotani, S Kuriki, H Karibe, N Nakasato
(eds), Recent Advances in Biomagnetism. Sendai:
Tohoku University Press, 129-32.
Schetzen M (1980):
The Volterra and Wiener Theories of Nonlinear Systems.
New York: John Wiley.
Sekihara K, Srikantan N, Poeppel D, Miyashita
Y (2000): Reconstructing spatio-temporal activities of
neuronal sources using MEG vector beamformer. Neuroimage 5: S485.
Sommerhoff G (1974). Logic
of the Living Brain. London: John Wiley.
Sporns O, Tononi G, Edelman GM (2000): Theoretical
neuroanatomy: relating anatomical and functional connectivity
in graphs and cortical connection matrices. Cereb
Cortex 10(2): 127-41.
Thatcher RW, Toro C, Pflieger ME, Hallett M
(1994): Human neuronal network dynamics using multimodal
registration of EEG, PET, and MRI. In: RW Thatcher,
M Hallett, T Zeffiro, ER John, M Huerta (eds), Functional
Neuroimaging: Technical Foundations.
San Diego: Academic Press, 269-78.
Valdes PA, Jiminez JC, Riera J, Biscay R, Ozaki
T (1999): Nonlinear EEG analysis based on a neuronal mass
model. Biol Cybern 81: 415-424.
Vazquez AL, Noll DC (1998): Nonlinear aspects
of the BOLD response in functional MRI. Neuroimage
7(2): 108-18.
von der Malsberg C, Singer W (1988): Principles
of cortical network organization. In: P Rakic and
W Singer (eds), Neurobiology of the Neocortex.
New York: John Wiley, 69-99.
Wagner M, Fuchs M, Wischmann HA, Köhler T,
Theißen A, Willemsen S (1999): fMRI-constrained source
reconstructions. In: T Yoshimoto, M Kotani, S Kuriki,
H Karibe, N Nakasato (eds), Recent Advances in Biomagnetism.
Sendai: Tohoku University Press, 137-40.
Wildgruber D, Erb M, Klose U, Grodd W (1997):
Sequential activation of supplementary motor area and
primary motor cortex during self-paced finger movement
in human evaluated by functional MRI. Neurosci Lett 227: 161-4.
Worsley JK (1994): Local maxima and the expected
Euler characteristic of excursion sets of {chi-square},
F and t fields. Advances in Applied Probability
26: 13-42.
Worsley KJ, Friston KJ (1995): Analysis of
fMRI time-series revisited – again. Neuroimage 2: 173-181.
Yates BJ, Stocker SD (1998): Integration of
somatic and visceral inputs by the brainstem: functional
considerations. Exp Brain Res 119: 269-75.
Zarahn E, Aguirre G, D’Esposito M (1997):
Empirical analyses of BOLD fMRI statistics. I. Spatially
unsmoothed data collected under null-hypothesis conditions.
Neuroimage 5: 179-97.

 Home
Current Issue
Table of Contents
Home
Current Issue
Table of Contents
