MGH-NMR
Center – Harvard Medical School Charlestown Massachusetts
USA
We compared the localization results between the two methods
and against the known source distributions. We quantified
the spatial and the temporal accuracy. The anatomic
selection of the sources also allowed us to examine the
effect of the source location on the results.
In the first part, we briefly present
the linear estimation and SPTF approaches and the generation
of the model data. We also describe the original method
we used to represent our localization results. Secondly,
we present the spatial and temporal accuracy of each method
for each simulation dataset which demonstrates the complementarily
of the two approaches.
1. Introduction
Using magnetoencephalography (MEG) one can make measurements
of brain activity with very high temporal resolution. The
localization of the underlying generators in the brain requires
solving the “inverse problem” of determining the
source distributions that give rise to some set of external
electromagnetic measurements. Numerous inverse algorithms
have been used to localize the activity, e.g. [Wood, C.C,
1985, Scherg, M 1992, Mosher, J.C. et Al, 1992, Hamalainen,
M., et al, 1993, George, J.S 1995]. Two promising approaches
are linear estimation [Hamalainen, M et al, 1993, Liu, A.K
et al,1998, Dale, A.M et al 1993, Wang, J.Z et al 1998,
Sarvas, J, 1987] and the Spatio-Temporal Fit (SPTF) [Schwartz,
D et al 1999]. Neither linear estimation nor SPTF require
any a priori information about the number of sources.
Both are user-independent methods that will give the same
localization regardless of the initial conditions.
The linear estimation approach computes an optimal linear
inverse operator, which explicitly minimizes the expected
error between the estimated and actual activity distribution.
The SPTF algorithm applies Principal Component Analysis
to the signal over a short moving window of time to build
a signal subspace and finally separates simultaneous asynchronous
dipolar sources.
The comparison of any two inverse methods is made more
complicated by the ill-posed nature of the inverse problem.
Theoretically, there are an infinite number of solutions,
which will reproduce the external measurements. Therefore,
with only external experimental measurements, it is not
possible to know with full accuracy the position and time
course of the actual generators. Electrophysiological
invasive recordings are essential to truly characterize
the internal source distributions. This work essentially
compares linear estimation and SPTF using simulated data,
allowing for comparisons against the “gold standard”
of the known source distributions.
Our model data was generated using realistic source distributions
constrained to the cortical surface with realistic time
courses of activation. The sources were placed in four
well-defined areas: frontal, precentral, temporal, and parietal.
We examined single and multiple sources with various amounts
of temporal overlap and temporal correlation.
We compared the localization results between the two methods
and against the known source distributions. We quantified
the spatial and the temporal accuracy. The simulated
sources always lied inside the gray matter of a realistic
head model, from one of our human subjects, with different
depths that allowed us to study the effect of various source
locations.
In the first part, we briefly present the linear estimation
and SPTF approaches and the generation of the model data.
We also describe the original method we used to represent
our localization results. Secondly, we present the spatial
and temporal accuracy of each method for each simulation
dataset.
2. Methods
2.1. Linear Estimation
The linear estimation approach [Hamalainen, M et al, 1993,
Liu, A.K et al,1998, Dale, A.M et al 1993, Wang, J.Z et
al 1998, Sarvas, J, 1987] computes an optimal linear inverse
operator (W), which maps the external electromagnetic
field measurements (x) into estimated source activities
( ) within the
brain. This inverse operator explicitly minimizes the expected
error between the estimated and actual activity distribution
(s). The calculation incorporates the forward solution
(also called the gain matrix (A)), spatial priors
(R) and sensor noise characteristics (C).
For these solutions, we constrained the source locations,
but not the source orientations, by the cortical surface
(i.e. anatomical constraint). The expression for the inverse
operator is:
) within the
brain. This inverse operator explicitly minimizes the expected
error between the estimated and actual activity distribution
(s). The calculation incorporates the forward solution
(also called the gain matrix (A)), spatial priors
(R) and sensor noise characteristics (C).
For these solutions, we constrained the source locations,
but not the source orientations, by the cortical surface
(i.e. anatomical constraint). The expression for the inverse
operator is:
 (1)
(1)
In these modeling studies, we assumed no prior spatial
information (R = I) and uncorrelated gaussian
noise with a signal-to-noise ratio of 10
(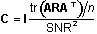 ).
).
This linear inverse operator provides an estimate of source
activity for each time point independently of the other
time points. The estimated source activities are given
by the simple expression:
 (2)
(2)
2.2. SPTF
The SPTF approach was extensively described and tested
in [Schwartz, D et al 1999]. We describe here the method
used to introduce anatomical constraints in the SPTF framework.
Classically the SPTF searches for an equivalent dipole
 to match, through a spatio-temporal analysis, the external electromagnetic
field measurements (x) with the computed field
to match, through a spatio-temporal analysis, the external electromagnetic
field measurements (x) with the computed field
 that is generated by the dipole. Let
that is generated by the dipole. Let
 be the
spatio-temporal matrix
be the
spatio-temporal matrix  containing the signals for
containing the signals for
 sensors at
sensors at  latencies. The SPTF performs a Principal Component Analysis (PCA) on the matrix
latencies. The SPTF performs a Principal Component Analysis (PCA) on the matrix

 .
The results are
.
The results are
 eigenvectors
eigenvectors
 and
and
 decreasing
decreasing
 eigenvalues. A discontinuity in the order of magnitude of the eigenvalues
eigenvalues. A discontinuity in the order of magnitude of the eigenvalues
 indicates the dimension of the signal subspace
indicates the dimension of the signal subspace
 of dimension
of dimension
 with
with
 .
.
 is then defined as follows:
is then defined as follows:

Figure 1. Source
time course used to model the temporal behavior of each
source
We computed the MEG signals generated by each sources individually
and by 3 combinations of 3 sources with differing amounts
of temporal overlap and temporal correlation between the
sources (See Table 1). For multiple source configurations,
each source had equal source power without equal sensor
power. One time point was generated every 10 ms, for 61
time points from 0 to 600 ms. To these signals, we added
gaussian noise with a signal-to-noise ratio (peak RMS) of
10.
TABLE 1. Temporal overlap and temporal
correlation for the two configurations of three sources
used in the simulations: Frontal, Parietal, Precentral and
Frontal, Precentral, Temporal.
| |
Temporal overlap and temporal correlation
|
Time courses
|
|
1
|
Temporal correlation = 0.0
|

|
|
2
|
Temporal overlap = 50 % to 100 %
Temporal correlation = 1.0
|

|
|
3
|
Temporal correlation = 0.75 to 1.0
|
|
2.4. Rendering
All results were represented on
the inflated cortical surface, which allows the visualization
of activity hidden in sulci. The localization of each dataset
results in a spatio-temporal movie. To give a more concise
representation of the data, we computed 1) the peak activation
and 2) the peak latency over the entire cortical surface.
1. Peak Activation: Maximum value (linear estimation
 or SPTF correlation)
found in each point of the cortical surface between 0 ms
and 600 ms. For both linear estimation and SPTF we used
a threshold determined automatically to represent only the
significant values.
or SPTF correlation)
found in each point of the cortical surface between 0 ms
and 600 ms. For both linear estimation and SPTF we used
a threshold determined automatically to represent only the
significant values.
2. Peak Latency: Latency of the peak activation
at each point of the cortical surface. The peak latency
was only represented when the corresponding peak activation
was greater than the automatically determined threshold.
We chose to represent peak latencies instead of onset latencies
because the peak latencies are insensitive to thresholding
effects. We used a rainbow color map (red, orange, yellow,
green, light blue, dark blue) to represent the peak latency.
2.5. Comparison of spatial and temporal accuracy
We compared both quantitatively
and qualitatively the localization results from linear estimation
and SPTF. For each the single source activation we evaluated
the localization accuracy by comparing the center of mass
of the actual source and center of mass of the localized
source. With multiple sources, it becomes problematic to
objectively define “sources.” Therefore, for
the multiple sources activation we evaluated the number
of sources effectively retrieved versus the actual number
of sources. Additionally we qualitatively compared the size
of the retrieved sources with the size of the actual sources
to evaluate the ability of each algorithm to separate these
sources. To evaluate the temporal accuracy, we computed
the mean and the standard deviation of the peak latency
at each actual source location for each data set.
3. Results
3.1. Single source activation
3.1.1. Spatial accuracy
TABLE 2. Distance between the Center of Mass of the localized
source and the Center of Mass of the actual source.
| |
Linear
Estimation
Distance (mm)
|
SPTF
|
| Frontal |
10.9
|
8.3
|
|
Temporal
|
15.3
|
11.1
|
|
Precentral
|
10.8
|
7.4
|
|
Parietal
|
13.7
|
14.4
|
The distances from the estimated center of mass to the
actual center of mass for the four different sources are
shown in Table 2. SPTF provides slightly better localization
than linear estimation, with an average improvement of 2.4
mm.
3.1.2. Temporal accuracy
TABLE 3. Peak Latencies for Single
Source.
| |
Linear EstimationPeak Latency (ms) |
SPTFPeak Latency (ms) |
Actual Peak Latency (ms |
| Frontal |
201 ± 7 |
224 ± 35 |
210 |
| Temporal |
200 ± 0 |
233 ± 12 |
210 |
| Precentral |
207 ± 4 |
198 ± 43 |
210 |
| Parietal |
201 ± 3 |
149 ± 43 |
210 |
The peak latencies for the four different source locations
are shown in Table 3. The actual peak latency is 210 ms.
Linear estimation provides an accurate and consistent estimate
of the peak latency. In contrast, SPTF has later peak latencies
(frontal and temporal) and earlier peak latencies (precentral
and parietal). In addition the variance of the peak latency
for SPTF is much larger.
3.1.3. Spatial extent versus Threshold

Figure 2. Spatial extent versus
threshold. Peak activation for three different thresholds.
We used a heat color map with transparent low values, the
mid values are represented in red, and the higher values
tend toward yellow. For the linear estimation the threshold
is in red (50%), full red and yellow correspond to 80% and
90% of the absolute peak activation, respectively.
For SPTF, threshold is in red (85%) and full red and yellow
correspond to 92% and 100% of the absolute peak activation,
respectively.
As expected (see Figure 2), raising the threshold
results in a more focal localization for both localization
approaches. Linear estimation is more sensitive to the threshold
selected. The parietal source is no longer visible when
using a threshold that results in sufficiently focal activity
of the precentral and frontal source. For SPTF, one can
see that a single threshold yields visible activity in all
three sources.
3.2. Multiple Sources
3.2.1. Spatio-temporal accuracy

Figure 3. Frontal, Parietal and
Precentral Source Localization (FPPR 3). Peak activation
for linear estimation (top) and SPTF (bottom) are shown
on the left. Peak latency for linear estimation (top)
and SPTF (bottom) are shown on the right. The MEG signals
are shown on the bottom.
Shown in Figure 3 are the localization results for
the Frontal, Parietal and Precentral sources for the third
temporal configuration. Consistent with the single
source results, the spatial localization of SPTF is more
focal than linear estimation. The frontal and precentral
activities are retrieved by both techniques. However,
the parietal activity is only recovered by SPTF. The
peak latencies are shown in Table 4. The temporal
agreement between the two methods is not as good as the
spatial agreement. The linear estimation peak latency,
for those sources that are well localized, is more accurate
and more consistent than the SPTF peak latencies.
TABLE 4. Peak Latencies for Multiple Extended
(2.0 cm diameter) Sources –
Frontal, Parietal and
Precentral. The Parietal source was not well localized
using linear estimation.
| Linear Estimation
Peak Latency (ms)
|
|
SPTF
Peak Latency (ms)
|
Actual latency (ms) |
| 1 |
|
|
|
| Frontal |
150
± 5 |
155
± 19 |
160 |
| Parietal |
358
± 23* |
316
± 28 |
360 |
| Precentral |
548
± 20 |
533
± 11 |
560 |
| 2 |
|
|
|
| Frontal |
150
± 5 |
52
± 5 |
160 |
| Parietal |
266
± 19* |
286
± 4 |
260 |
| Precentral |
352
± 4 |
363
± 29 |
360 |
| 3 |
|
|
|
| Frontal |
151
± 5 |
56
± 9 |
160 |
| Parietal |
196
± 20* |
205
± 0 |
210 |
| Precentral |
250
± 0 |
271
± 20 |
260 |
|
|
|
|
3.2.2. Linear estimation

Figure 4. Frontal, Precentral and
Temporal Extended Source Localization (FPRT 2). Peak activation
for linear estimation (top) and SPTF (bottom) are shown
on the left. Peak latency for linear estimation (top)
and SPTF (bottom) are shown on the right. The MEG signals
are shown on the bottom.
TABLE 5.
Peak Latencies for Multiple Extended (2.0 cm diameter)
Sources – Frontal, Precentral and Temporal. * The
Temporal source was not well localized using linear estimation.
|
Linear
Estimation
Peak Latency (ms)
|
SPTF
Peak Latency (ms)
|
Actual
Latency (ms)
|
|
1
|
|
|
|
|
Frontal
|
150
± 5
|
161
± 24
|
160
|
|
Precentral
|
357
± 4
|
392
± 18
|
360
|
|
Temporal
|
517
± 49*
|
536
± 27
|
560
|
|
2
|
|
|
|
|
Frontal
|
151
± 5
|
54
± 8
|
160
|
|
Precentral
|
250
± 0
|
252
± 36
|
260
|
|
Temporal
|
340
± 25*
|
385
± 2
|
360
|
|
3
|
|
|
|
|
Frontal
|
151
± 5
|
96
± 57
|
160
|
|
Precentral
|
208
± 3
|
220
± 38
|
210
|
|
Temporal
|
221
± 34*
|
262
± 12
|
260
|
Shown in Figure 4 are the localization results for
Frontal, Precentral and Temporal extended sources with different
amount of temporal overlap (FPPRT 1, FPRT 2 and FPRT 3).
These results are similar to those of FPPR. The spatial
localization of SPTF is more focal than the one using the
linear estimation. For both techniques, the frontal
and precentral sources are well localized, however the temporal
source is poorly localized. The peak latencies for both
techniques are shown in Table 5.
4. Discussion
We found little variation in the spatial localization as
a function of source extent for both techniques. This
essentially reflects the poor spatial resolution of the
forward operator. That is, nearby locations in the
brain generate very similar forward solutions. Subsequently,
those locations are difficult to separate. If one
has a priori information that suggests the sources
are indeed focal, single and multiple dipole approaches
may be more appropriate than either linear estimation or
SPTF.
The center of mass of activation was slightly better for
SPTF than linear estimation. Comparison of the spatial
extent of activation between the two techniques shows that
SPTF is consistently more focal than linear estimation.
Since SPTF performs the localization, not on the actual
signal, but on a reconstructed signal subspace generated
from a moving time window, those localizations become more
robust in presence of noise. This allows a more aggressive
thresholding of the estimated probability of activation.
In addition, small amplitude sources are better reconstructed
using SPTF than linear estimation because the probability
may still remain high even while the absolute activation
is small compared to other sources.
The entire issue of thresholding is, however, a difficult
one, and altering the threshold in any localization technique
will alter the spatial extent of the localization. We addressed
this issue using an automatic thresholding approach for
both methods. This gives us an objective way to compare
the results from linear estimation and SPTF. For a single
source, it is always possible to generate a more focal solution
by increasing the threshold. When there are multiple sources
with equal source power but not necessarily equal sensor
power, altering the threshold can result in variable spatial
localizations where entire sources may be lost at higher
and more focal thresholds. In physiological measurements,
one would expect comparable source power rather than sensor
power. The sensor power for a given source is determined
by the local cortical morphology.
In comparison to the spatial localization, the temporal
information given by linear estimation is more consistent
and, in some cases, more accurate than SPTF. Since the
linear estimation approach works directly with the measurement
data, no temporal dynamics of well-reconstructed sources
are lost. For SPTF, the estimated probability does not
necessarily correlate to the actual intensity of the source.
Subsequently, the maximum of probability does not always
occur at the actual peak activation.
In the multiple source conditions, the parietal and temporal
sources were difficult to localize, while the frontal and
precentral were always well reconstructed. Alone,
both the parietal and temporal sources were well retrieved.
This variability in localization accuracy demonstrates the
effect of the sensor sensitivity to different locations
in the brain. For a given activity, the sensor power
(i.e. norm of the gain vector) is smaller for the parietal
and temporal sources than for the frontal and precentral.
As a result, when several sources are active, the frontal
and precentral sources will dominate the measurements.
This discrepancy in sensor power will also result in lower
signal to noise ratios for the parietal and temporal sources,
resulting in poor localization accuracy in these anatomic
regions. This effect is likely the cause of the poor
localizations in the temporal lobe as reported by [25].
The effect of sensor power is greater in the linear estimation
approach, since a low threshold results in additional spurious
activity for our parietal and temporal sources. Hence,
with real data, it is not possible to differentiate “noise
activity” from weak sources.
Linear estimation gives a more consistent temporal localization
than SPTF. The temporal discrepancies between SPTF and linear
estimation are higher than the ones seen for single source
activation. For multiple sources activation the estimated
probability computed by SPTF is higher when the source is
activated alone and decreases when additional sources appear
in the signal. Thus the estimated peak latency usually occurs
earlier than the actual peak. This is especially obvious
for the SPTF estimate of the frontal source whose peak latency
occurs at 50 ms, well before the actual peak at 160 ms.
One of the theoretical drawbacks of SPTF is its sensitivity
to correlated sources. SPTF looks for single dipolar activity
in the signal subspace and thus is not able to separate
multiple correlated sources. We expected the SPTF results
to be worst for FPPR 3 and FPRT 2, which had the greatest
correlation between sources. In addition, we expected the
linear estimation results to be insensitive to correlation.
Not surprisingly, the linear estimation localizations were
essentially the same for all levels of correlation. However,
the SPTF spatial localizations remained very accurate even
for FPPR 3 and FPRT 2. This somewhat unexpected result is
due to the use of the anatomic constraint. When two sources
are highly correlated the SPTF maximum probabilities would
be in some other location beside the actual sources. This
maximum typically would not be on the cortical surface while
secondary peaks will still be found at the actual source
locations.
5. Conclusion
Our results demonstrate the complementary
strengths of linear estimation and SPTF. While both linear
estimation and SPTF provide comparably accurate spatial
localization of single and multiple extended sources, SPTF
gives more focal solutions. Surprisingly, SPTF is not very
sensitive, in terms of spatial accuracy, to the temporal
correlation between the sources. Linear estimation provides
more accurate temporal information especially when several
sources are simultaneously activated. We can exploit the
superior spatial accuracy of SPTF by incorporating the SPTF
spatial localizations as a prior in the linear estimation
approach. In future modeling and experimental work we will
explore and utilize the greater spatio-temporal accuracy
of this combined linear estimation – SPTF technique.
Acknowledgement
We thank Dr. Anders Dale at the
MGH-NMR Center, Mass. General Hospital for helpful discussion
and support.
6. References
Barth, D.S. and S. Di, Three-Dimensional analysis of
auditory-evoked potentials in rat neocortex. J of neurophysiol,
1990: p. 1527-1536.
Dale, A.M., B. Fischl, and M.I. Sereno, Cortical Surface-Based
Analysis I: Segmentation and Surface Reconstruction.
NeuroImage, 1999. 9: p. 179-194.
Dale, A.M. and M.I. Sereno, Improved localization of
cortical activity by combining EEG and MEG with MRI cortical
surface reconstruction: A linear approach. J cog neurosci,
1993. 5(2): p. 162-176.
Fischl, B., M.I. Sereno, and A.M. Dale, Cortical Surface-Based
Analysis II: Inflation, Flattening, and a Surface-Based
Coordinate System. NeuroImage, 1999. 9: p. 195-207.
George, J.S., et al., Mapping function in the
human brain with MEG, anatomical MRI and functional MRI.
J clin neurophys, 1995. 12(5): p. 406-431.
Hamalainen, M.S. and J. Sarvas, Realistic conductivity
geometry model of the human head for interpretation of neuromagnetic
data. IEEE trans biomed eng, 1989. 36: p. 165-171.
Hamalainen, M., et al., Magnetoencephalography
- theory, instrumentation, and application to noninvasive
studies of the working human brain. Review of modern
physics, 1993. 65: p. 413-497.
Kaga, K., Hink, R.F., Shinoda, Y., Suzuki, J., Evidence
for a primary cortical origin of a middle latency auditory
evoked potential in cats. Electroenceph clin neurophysiol,
1980. 50: p. 254-266.
Knuutila, J.E.T., et al., A 122-channel whole-cortex
squid system for measuring the brains magnetic fields.
IEEE trans magn, 1993. 29(6): p. 3315-3320.
Liegois-Chauvel, C., et al., Evoked potentials
recorded from the auditory cortex in man: evaluation and
topography of the middle latency components. Electroenceph
clin neurophysiol, 1004. 92: p. 204-214.
Liu, A.K., J.W. Belliveau, and A.M. Dale, Spatiotemporal
imaging of human brain activity using functional MRI constrained
magnetoencephalography data: Monte Carlo simulations.
Proc natl acad sci USA, 1998. 95(15): p. 8945-50.
Mikuni, N., et al., Simultaneous recording of
epileptiform discharges by MEG and subdural electrodes in
temporal lobe epilepsy. Neuroimage, 1997. 5(4
Pt 1): p. 298-306.
Meijs, J.W.H., et al., On the numerical accuracy
of the boundary element method. IEEE trans biomed eng,
1989. 36(10): p. 1038-1049.
Meijs, J.W.H., et al., On the magnetic field
distribution generated by a dipolar current source situated
in a realistically shaped compartment model of the head.
Electroenceph clin neurophysiol, 1987. 66: p. 286-298.
Mosher, J.C., P.S. Lewis, and R.M. Leahy, Multiple
dipole modeling and localization from spatio-temporal MEG
data. IEEE trans biomed eng, 1992. 39(6): p.
541-557.
de Munck, J.C., A linear discretization of the volume
conductor boundary integral equation using analytically
integrated elements. IEEE trans biomed eng, 1992. 39(9):
p. 986-990.
Oostendorp, T.F. and A. van Oosterom, Source parameter
estimation using realistic geometry in bioelectricity and
biomagnetism, in Biomagnetic Localization and 3D
Modeling, Report TKK-F-A689, J. Nenonen, H.M. Rajala,
and T. Katila, Editors. 1992, Helsinki University of Technology:
Helsinki.
Sarvas, J., Basic mathematical and electromagnetic
concepts of the biomagnetic inverse problem. Phys med
biol, 1987. 32(1): p. 11-22.
Schwartz, D., et al., Evaluation of a new MEG-EEG
spatio-temporal localization approach using realistic sources.
Brain topography, 1999: p. In Press.
Scherg, M., Fundamentals of dipole source potential
analysis, in Auditory evoked magnetic fields and
electric potentials, F. Grandori, M. Hoke, and G.L.
Romani, Editors. 1990, Karger: Basel. p. 40-69.
Scherg, M., Functional imaging and localization of
electromagnetic brain activity. Brain topography, 1992.
5(2): p. 103-11.
Smith, D.I.a.K., N., Intracranial and extracranial
recordings of the auditory middle latency response.
Electroencephlogr clin neurophysiol, 1987. 71: p.
293-303.
Steinschneider, M., Tenke, C.E., Schroeder, C.E., Javitt,
D.C., Simpson, G.V. Arezzo, J.C., Vaughan H.G. Jr, Cellular
generators of the cortical auditory evoked potential initial
component. Electroenceph clin neurophysiol, 1991. 84:
p. 196-200.
Wang, J.Z., S.J. Williamson, and L. Kaufman, Magnetic
source images determined by a lead-field analysis: the unique
minimum-norm least-squares estimation. IEEE trans biomed
eng, 1992. 39(7): p. 665-75.
Wood, C.C., et al., "Electrical sources in
human somatosensory cortex: Identification by combined magnetic
and potential recordings." Science, 1985. 227:
p. 1051-1053.

 Home
Current Issue
Table of Contents
Home
Current Issue
Table of Contents


