 |
International Journal of Bioelectromagnetism Vol. 5, No. 1, pp. 185-186, 2003. |
www.ijbem.org |
|
A Model for the Passive Cardiac Conductivity Jeroen G. Stinstra, Bruce Hopenfeld,
and Rob S. MacLeod CardioVascular Research and Training Institute, and Bioengineering
Department, University of Utah, Correspondence: Jeroen Stinstra, CVRTI, University of
Utah, Bldg 500, Room 207, 95 South, 2000 East, 84112-UT, Salt Lake City, USA.
Abstract. Since the reported values
of the cardiac bidomain conductivities vary over a wide range, we decided
to estimate these conductivities based on a computer model of the underlying
tissue morphology. This model was adapted to compute the four bidomain conductivity
values (the "bulk" conductivities along and across the fiber-direction
both intracellular and extracellular). The model consisted of two conductive
domains, the intracellular space and the extracellular space, separated by
the highly resistive cell membrane. We modeled a cluster of 125 cells and
added details like the capillary system. We varied certain parameters (e.g.
cell length) to estimate their effect on the bidomain conductivities.
Keywords: Cardiac Tissue; Bidomain Model; Cardiac Electrical Conductivity 1. Introduction The passive electrical conductivity of cardiac tissue is an important parameter of computer models that attempt to emulate various aspects of cardiac electrophysiology. A nice example of this influence is given by Johnston [Johnston, 2002], who estimated epicardial potentials based on three different sets of bidomain conductivity values commonly cited in literature. At least two of the sets of conductivity values led to significantly different epicardial potential distributions. Thus, for accurate whole-heart electrical modeling, it may be important to have accurate bidomain conductivities. However, reported myocardial conductivity values, based upon experimental data, vary over a wide range. The uncertainty in measured conductivities is high because the measurements are very complicated and it is difficult to extract the bidomain conductivities from the experimental data. For instance, if the measurements are carried out in vitro, the accuracy may be low because tissue properties change rapidly outside the body, because the tissue is deprived of blood. If the measurements are carried out in vivo, animal tissue is often used instead of human tissue. Another problem is that the effective conductivity cannot be determined unambiguously, as the conductivity is dependent on the direction, the uniformity, the frequency, and the strength of the applied current. One way to avoid some of these difficulties is to model the conductive process at a cellular scale and to extrapolate, from this model, estimates of the effective properties at a tissue or organ scale. Such small scale models may help to estimate upper and lower bounds for the bidomain conductivities. In particular, this paper will provide estimates of the bidomain conductivities as a function of tissue specific characteristics, i.e. tissue morphology and the resistances of the cellular components such as gap junctions. 2. Material and Methods In order to predict the passive effective conductivity of cardiac tissue, we created geometrical models of cardiac tissue. The model describes cardiac tissue as consisting of an extracellular volume and an intracellular volume separated by a poorly conducting cell membrane. Both the intracellular and extracellular volume was considered to consist of a substance that had a homogeneous, ohmic conductivity. The ohmic effective conductivity of the tissue was computed by assuming an externally applied electric field and computing the resulting current density in the tissue model. The "effective conductivity" was equal to the computed net current divided by the voltage drop across the tissue. The effective conductivity thus assumed a homogeneous medium but not anisotropic medium. By applying the external electric field in different directions we could compute the anisotropy factor of the tissue. In particular, we applied the field along the fiber direction and perpendicular to the fiber direction in order to compute the effective conductivity in both directions. The effective conductivity was split further into the effective intracellular and effective extracellular conductivity by assuming that the cell membrane was not conducting. This situation resembles real world experiments that employ injected current and make measurements reasonably far away from both the current source the current sink. In these conditions, the volume currents have spread evenly between both the intra- and extracellular spaces. We computed both the intra- and extracellular conductivities as averages over an entire volume of tissue.
 Figure 1. An example of a cluster of myocytes in the model. Each myocyte is irregularly shaped, though in such a way that they all fit together like a jig-saw puzzle. This illustration shows only the cells that form the intracellular space. The extracellular space is carved around these cells. In order to estimate the conductivity as accurately, as possible, we created a program that generated a cluster of irregular-shaped myocytes (see Fig. 1). The shapes were chosen in such a way that the average length and average cross-section of a myocyte equaled the values found in the literature. Further, we assumed that the intracellular specific conductivity was 0.3 S/m and the extracellular conductivity was 1.8 S/m. The gap junctions connecting the myocytes were assumed to be at the end-planes of the myocytes (in the planes perpendicular to the fiber direction). The resistance of the gap-junctions between two neighboring myocytes, with an overlap in cross-section of about half the average cross-section of a myocyte, was 2.5 MΩ. The overall tissue volume fraction occupied by the myocytes was 85 percent, the volume fraction occupied by the interstitial space was 10 percent, and the remaining 5 percent was occupied by a non-conducting capillary system. 3. Results
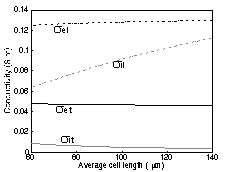 Figure 2. The estimated effective "bidomain" conductivities as a function of the myocyte length. These values were computed using a realistically shaped model of a cluster of 125 myocytes. Here, σel and σet are the effective extracellular conductivities in the longitudinal (along the fibers) and transverse directions, and σil and σit are the effective intracellular conductivities. Fig 2. shows the influence of the average cell length on the effective bulk conductivities. We found that the extracellular conductivities are hardly influenced by the cell length, whereas the intracellular conductivities depend on this parameter. The figure also shows that the anisotropy ratio for the intracellular conductivities (ranging between 10 and 25) significantly differs from the extracellular one (a value of 2.5). 4. Discussion Although many investigators consider the anisotropy ratios of the intra and extracellular spaces to be equal, the results of this realistically-shaped model seem to indicate otherwise. Not only did both anisotropy ratios differ, the intracellular one also depended on the actual length of the cells. In the near future, we hope to combine the results of this model with diffusion-tensor MR images and information regarding the actual ultrastructure throughout the heart in order to create a more realistic model of cardiac conductivity throughout the heart. An additional goal is to estimate the effects of ischemia on the bidomain conductivities and to estimate the resulting changes in epicardial and body surface potentials. References Johnston PR. Tissue conductivity and ST depression in a cylindrical left ventricle. Int J Bioelectromagnetism, 4(2), 61-62 2002.
© International Society for Bioelectromagnetism
|

