 |
International Journal of Bioelectromagnetism Vol. 5, No. 1, pp. 158-159, 2003. |
www.ijbem.org |
|
Gap Junction Remodeling and Alterations of Conduction Properties in Hypertrophied Cardiac Muscle Haruo Honjoa, Mahmud
Uzzamana, Luni Emdada, Chieko Sasanoa,Yoshiko
Takagishia, aResearch Institute of Environmental Medicine,
Nagoya University, Nagoya, Japan Correspondence: H Honjo, Department of Humoral Regulation,
Research Institute of Environmental Medicine, Nagoya University, Abstract. Normal action potential
propagation in the heart depends on electrical coupling through gap junctions.
In rat models of pressure overload-induced ventricular hypertrophy, we found
characteristic changes of gap junction distribution; immunolabeled connexin43
(Cx43) was redistributed from the intercalated disks to lateral cell borders
and the density of Cx43 in the intercalated disks was decreased in hypertrophied
ventricles. Extracellular potential mapping revealed that, in hypertrophied
ventricles, conduction velocity was decreased in a direction parallel to the
fiber orientation whereas unaffected in a direction across the fiber direction.
These results suggest that remodeling of gap junction distribution may underlie
alterations of anisotropic conduction properties of the hypertrophied ventricle.
Keywords: Gap Junction; Hypertrophy; Anisotropic Conduction Property 1. Introduction Cardiac hypertrophy is known to increase the risk of ventricular tachyarrhythmias, pump failure and sudden cardiac death. Alterations in both active and passive membrane properties may contribute to the arrhythmogenic substrates in the hypertrophied ventricle. Gap junctions constitute a major determinant of the passive membrane properties and changes in gap junctions could be a cause for abnormal conduction properties. In this study, we investigated changes in connexin43 (Cx43), the major myocardial gap junction protein, distribution and alterations of anisotropic conduction properties in pressure overload-induced ventricular hypertrophy in rats [Uzzaman et al., 2000; Emdad et al., 2001]. 2. Material and Methods Injection of monocrotaline (MCT, 60 mg/kg), which is known to cause pulmonary hypertension, to rats resulted in the development of right ventricular (RV) hypertrophy 4 weeks after the injection. LV hypertrophy was produced in rats by banding of the abdominal aorta (8-12 after the surgery). Single ventricular myocytes and cryosections of ventricular tissues were prepared and they were immunolabeled with an anti-Cx43 antibody (Chemicon). The samples were viewed using a confocal microscope. Ultrastructure of cell-to-cell junctions was examined by transmission electron microscopy. The conduction of excitation in the epicardial surface of hypertrophied RV tissues from MCT-treated rats was assessed by high-resolution mapping of extracellular potentials during point stimulation. 3. Results 3.1. Altered Cx43 Gap Junction Distribution in Hypertrophied Ventricles In normal ventricular cells, Cx43 gap junctions were largely confined to the intercalated disks at the cell termini (Fig. 1A). Hypertrophied RV cells of MCT-treated rats (Fig. 1B) and hypertrophied LV cells of aorta-banded rats (Fig. 1C) showed dispersion of Cx43 labeling over the entire cell surface rather than localization in the intercalated disk area. Similar changes in the gap junction distribution were seen in the ventricular tissues sectioned longitudinally (parallel to the fiber orientation).
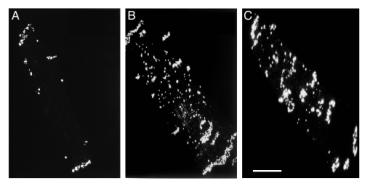 Figure 1. Immunolocalization of gap junctions in isolated ventricular myocytes. Confocal projection images constructed from optical section through the full thickness of myocytes labeled for Cx43. A, B, RV myocytes from a control (A) and a MCT-treated (B) rats; C, LV myocyte from an aorta-banded rat. Scale bar, 20 µm. En-face views of intercalated disks of immunolabeled normal ventricular tissues showed a characteristic pattern of gap junction distribution with small immunolabeling spots surrounded by large spots at the disk periphery. The former domains are small discoid gap junctions in the plicate region (the adhesive component of the disk structure) and the latter ones represent large ribbon-like gap junctions. In hypertrophied myocardium, there was a loss of the central small gap junctions giving rise to a significant reduction in the proportion of the disk occupied by gap junctions (by 25-30 %). 3.2. Ultrastructure of Remodeled Gap Junctions In the normal ventricular myocardium, gap junctions were recognized in longitudinally orientated (interplicate) segments as well as in the transverse (plicate) segments of the intercalated disks. In the hypertrophied ventricle, gap junctions in the plicate segment were less abundant than in controls. This corresponds to the immunolabeling observation showing a loss of centrally located small Cx43 positive spots in the intercalated disk viewed on face. In addition, gap junctions were frequently seen at sites distant from the intercalated disks. These included gap junctions forming side-to-side contact of myocytes and those showing annular profiles in the cytoplasm. The presence of the latter structure may indicate increased internalization of gap junctions during remodeling. 3.3. Anisotropic Conduction Properties In the normal RV myocardium, the epicardial activation pattern showed normal uniform anisotropy with higher conduction velocity parallel to the myocardial fiber orientation (θL) than that across it (θT). In the hypertrophied RV of MCT-treated rats, θL was significantly less (by 30 %) than controls, but there was no significant change in θT. The anisotropic ratio of conduction velocity (θL/θT) was significantly lower in the MCT-treated rats (1.38 in average) than controls (1.98). 4. Discussion In this study with rat models, we found that ventricular hypertrophy induced by pressure overload was associated with marked disorganization of Cx43 gap junction distribution: in hypertrophied ventricles, Cx43 gap junctions were dispersed over the cell surface rather than confined to the intercalated disk area and there was a significant decrease in the disk gap junction density. Some of these displaced gap junctions at sites of distance away from the intercalated disks formed side-to-side contacts but others were internalized gap junction structures presumably without function (annular gap junctions) [Uzzaman et al., 2000; Emdad et al., 2001]. Notwithstanding the need for caution in relating Cx43 distribution to electrophysiological properties, one predicted effect of the decreased gap junction density in the intercalated disk might be to reduce local current flow and thus conduction velocity parallel to the myocardial fiver orientation. In hypertrophied RV myocardium, in fact, the longitudinal conduction velocity was significantly decreased but the transverse conduction velocity was unaffected, giving rise to a significant reduction of anisotropic ratio compared with controls. References Emdad L, Uzzaman M, Takagishi Y, Honjo H, Uchida T, Severs NJ, Kodama I, Murata Y. Gap junction remodeling in hypertrophied ventricles of aortic-banded rats: prevention by angiotentin II type 1 receptor blockade. Journal of Molecular and Cellular Cardiology, 33: 219-231, 2001. Uzzaman M, Honjo H, Takagishi Y, Emdad L, Magee AI, Severs NJ, Kodama I. Remodeling of gap junctional coupling in hypertrophied right ventricles of rats with monocrotaline-induced pulmonary hypertension. Circulation Research, 86: 871-878, 2000.
© International Society for Bioelectromagnetism
|

