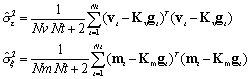The Log-posterior probability distribution of all the parameters
when the data is given, takes the form:


Where ,
,
 ,
,
 and
and
 are the elements of
are the elements of
 , the diagonal of
, the diagonal of
 ,
,
 and
and

 , respectively,
with i = 1..Ng.
, respectively,
with i = 1..Ng.
An intuitive interpretation of these estimators is that activity
is estimated as a linear combination of PCD and NMD neuroimages,
and in turn, influences their estimation. Note that the
estimation of all hyperparameters comes from the data itself.
4. Simulation Results
In this section we report the results of comparing measures
of quality of the Tomographic methods derived from the Pont
Spread Function (PSF) of Low Resolution Electric Tomography
(LORETA, [Pascual-Marqui1994]), EEG  fMRI, and fMRIEEG. This was carried out for the
grid of points inside the head that represents the possible
generators (i.e. gray matter) for which the PCD is computed.
In the simulations the same spatial resolution for EEG/MEG
Inverse Problem, and fMRI is assumed, and a grid of 3433
points equally spaced inside the gray matter is constructed,
matching the spatial locations of the voxels in the fMRI.
The simulation consisted in generating the EEG/MEG and fMRI
data, using the forward problem equations (1) and (6). The
PCD and NMD used correspond to the activation of a single
unit magnitude dipole located in each point of the grid
with a Gaussian variation in time (Figure
1). This curve was discretized at five time points were
simulated. A measurement noise level of 10% of the peak-to-peak
voltage/fMRI signal is assumed in the computations. The
PSF in each point, was computed at the time instant where
the signal to noise ratio is maximum in both EEG/MEG and
fMRI. All measurement units have been omitted for the sake
of simplicity.
fMRI, and fMRIEEG. This was carried out for the
grid of points inside the head that represents the possible
generators (i.e. gray matter) for which the PCD is computed.
In the simulations the same spatial resolution for EEG/MEG
Inverse Problem, and fMRI is assumed, and a grid of 3433
points equally spaced inside the gray matter is constructed,
matching the spatial locations of the voxels in the fMRI.
The simulation consisted in generating the EEG/MEG and fMRI
data, using the forward problem equations (1) and (6). The
PCD and NMD used correspond to the activation of a single
unit magnitude dipole located in each point of the grid
with a Gaussian variation in time (Figure
1). This curve was discretized at five time points were
simulated. A measurement noise level of 10% of the peak-to-peak
voltage/fMRI signal is assumed in the computations. The
PSF in each point, was computed at the time instant where
the signal to noise ratio is maximum in both EEG/MEG and
fMRI. All measurement units have been omitted for the sake
of simplicity.
Based on the PSF for both EEG
fMRI and LORETA, some quality measures for characterizing
electrophysiological Tomography techniques was computed,
namely, the Localization Error (LE), Full Width at Half
Maximum (FWHM) of the PSF, and visibility. The definition
used for LE was that described in [Pascual-Marqui1994]:
the distance between the location of the simulated dipole,
and the maximum of the estimated PCD, We define the FWHM
directly from the definition by using a nonparametric estimation
of the PSF. The measure of visibility was defined as the
maximum amplitude of the PSF. The density estimator used
was the Naradaya-Watson estimator [Stone 1977].
The measures of quality are summarized in form of nonparametric
regression curves in Figures
2-4. Additionally nonparametric standard errors are
also included in the figures. Nonparametric regressions
were carried out also using the NW estimator.
Figure
2 shows the LE. For LORETA this error decreases monotonically
with the eccentricity of the source, from 25 to 14 mm, while
for EEG
fMRI the LE is constant and almost zero for all points.
It is clear that the spatial information offered by the
fMRI measurements plays an important role in the improvement
of the LE.
Figure
3 shows the FWHM. There is a significant reduction of
the FWHM for EEG
fMRI with respect to LORETA for all points, indicating an
increased spatial sensitivity.
One
difficulty of linear inverse methods is the poor visibility
of deep sources, which are masked by those sources closer
to the sensors. Figure
4. Illustrates that the visibility of LORETA increases
exponentially with the eccentricity with a maximum value
of approximately 10-2. In contrast EEG
fMRI shows an almost constant behavior, near the optimal
value of 1.
In
addition to the results of regression equations for all
grid points, it is illustrative to compare the actual tomographic
solutions for selected grid points. In this case the results
of LORETA and EEG
fMRI are compared with a fMRI constrained minimum norm type
electrophysiological solution (fMRIEEG) previously described in the
literature ([Dale 1993;Dale2000]).
In
Figure 5-11
shows the results for a source simulated to be in the temporal
region. Figure
5 shows the actual source. In Figure
6-8 the temporal evolution of the activation, the modulus
of the NMD and the PCD are shown as estimated with EEG
fMRI are shown. The modulus of the PCD is reconstructed
with a small decrease in the amplitude of the simulated
dipole (see fig. 1), which is consistent with the small
decay of the visibility with the distance to the sensors.
In the case of the hemodynamic variable, the activation
is slightly overestimated. Figure
9 shows the map of the PCD obtained with LORETA showing
a wide region of activation, and a significant bias in the
spatial location of the maximum activity that is due to
the smoothing properties of the technique. When using fMRIEEG method (fig. 10) a peak at
the actual location of the simulated dipole is observed
probably an inheritance of the minimum norm part of the
method. The estimation by EEG ↔ fMRI (Fig. 11) is almost indistinguishable
from the ideal PCD used for the simulations.
It
was also thought convenient to explore the results of a
simulation using a distributed source. Therefore a Gaussian
function was placed in the occipital region as the simulated
source (Fig. 12). The PCD estimated by LORETA is shown in
Fig. 13. As expected, LORETA performs very well in this
case though there is a small error in the localization error
and a reduced visibility. The solution for fMRIEEG method is shown in Fig 14.
The reconstruction is quite poor, with maximal activation
at the points nearest to the sensors. A sharp frontier for
the estimated source is also shown showing that this method
is excessively biased for those points where is fMRI activity.
For the EEG/MEG
fMRI model, there is also a good reconstruction of the spatial
distribution of the activity (fig 15), and, unlike LORETA,
there is no localization error of the maximum. Nevertheless,
there is a significant bias in the estimation of the amplitude
of the source.
A quantitative comparison of the simulation results is
summarized in tables 1-2.
5. Analysis of a Somatosensory Experiment
The description of the somatosensory experiment analyzed
here is described fully in [Hoechstetter 2000].
Anatomical MRI: A T1 anatomical image was image
was obtained and used for registration of MEG and fMRI procedures.
The scalp and cortex were extracted using software developed
at the Montreal Neurological Institute by MacDonald [MacDonald
et al. 2000] resulting in triangulated surfaces with
normal vectors.
MEG data: Somatosensory evoked fields were recorded
from a healthy adult using a Neuromag-122 whole head MEG
system with 118 valid sensors used. Prior to recording head
position was determined by four coils attached to the scalp.
These served to transform the sensor positions and orientations
into the MRI space. A 3 sphere model was fitted to the head
using the scalp surface.
Brief tactile pressure pulses were delivered by finger
clips (BTI) at a constant inter-stimulus interval of 1.03
s. to the tip of the left index finder. Data were sampled
at 769 Hz. Event Related Magnetic Fields (ERF) to the SS
stimuli were obtained from 739 individual trials, yielding
a very good signal to noise ratio. The time course of the
ERF was assessed by means of a statistical analogue of the
Global Field Power recently described by [F.Carbonell 2001]
(consisting of a Hotelling’s T2 statistic over the
MEG sensors evaluated at each time point) in order to test
the presence of a mean vector. The composite alpha level
for the whole time epoch is derived from the theory of random
fields [Worsley 1994]. The results fo this computation are
shown in Figure
16. As can be seen 3 major peaks are present and these
were selected for further processing. In other words the
magnetic data to be processed consisted of 3 vectors  .
.
Sources were assumed to be restricted to a 2048-point grid
on the cortical surface as defined by the cortical triangulation.
The magnetic lead field was obtained using the sensor position
and source positions and orientations. Figure
17-a shows the SPM of the LORETA solution for the first
vector of the MEG data.
fMRI data: A total of 190 frames where gathered,
one every 2 sec. Two conditions were gathered in a blocked
design: Baseline vs. Activated state, lasting 20 seconds
each. The stimulus was the same as for the MEG recording.
Ten slices were selected to pass through the region of interest.
The activity in cortex was estimated by an interpolation
procedure, sampling restricted to the same points used for
the cortical grid described in the previous section. Figure
17-b shows the surface SPM map for this data.
Results of applying the EEG
fMRI fusion method are shown in the SPM map in Figure
17-c. As can be seen the localization of activity is
much more focal than for LORETA.
6. Final Considerations
The results presented seem to indicate significant advantages
for electrophysiological and fMRI image fusion.
The simulation results are very promising. It should be
taken into consideration that the cortical PSF based on
optical measurements has been estimated in primary visual
cortex of the macaque to be 1.5 mm (antero-posterior) by
2.7 mm (medio lateral) ([Grinvald 1994]), while the fMRI
resolution in human visual cortex has been estimated to
be 3.5 mm ([Engel 1997]). EEG
fMRI preserves the good spatial resolution of the imaging
techniques mentioned before, with an average FWHM of the
order of 1.4 mm. This is much better performance than that
of other techniques. What is more important there are considerable
improvements in visibility which seems to be the weakness
of linear inverse solutions.
The preliminary results presented for the somatosensory
data show that the methods proposed may be used for the
analysis of actual data, a concern with many current Bayesian
image analysis methods.
There is a number of directions in which the present work
is being extended. One is concerned with the introduction
of temporal correlations in the a priori information. More
importantly, the models expressed above are valid for a
single experimental condition. This is not the most frequent
situation either in fMRI or electrophysiology in which either
event related data or block experiments are gathered under
some experimental design. The extension of Statistical Parametric
Mapping methods for the fusion model is a natural development
of the Bayesian approach presented here which will be subject
of subsequent presentations.
7. Acknowledgements
We wish to thank Prof. Michael Scherg for providing the
experimental data analyzed in Section
References
Ahlfors, S. P. Simpson G. V. Dale A. M. Belliveau J. W.
Liu A. K. Korvenoja A. Virtanen J. Huotilainen M. Tootell
R. B. Aronen H. J. & Ilmoniemi R. J. Spatiotemporal
activity of a cortical network for processing visual motion
revealed by MEG and fMRI. Neurophysiology 82, 2545-2555.
1999.
Belliveau, J. W. Kennedy D. N. McKinstry R. C. Buchbinder
B. R. Weisskoff R. M. Cohen M. S. Vevea J. M. Brady T. J.
& Rosen B. R. Functional mapping of the of the humn
visual cortex by magnetic resonance imaging. Science
254, 716-719. 1991.
Dale, A. M. Lio A. K. Fischl B. R. Buckner R. L. Belliveau
J. W. Lewine J. D. & Halgren E. Dynamic statistical
parametric mapping: combining fMRI and MEG to produce
high-resolution spatiotemporal maps of cortical activity.
Neuron 26, 55-67. 2000.
Dale, A. M. and Sereno M. I. Improved localization of ciortical
activity by combining EEG and MEG with fMRI cortical surface
reconstruction: a linear approach. J.Cogn.Neurosci.
5, 162-176. 1993.
Engel, S. A. Glover G. H. and Wandell B. A. Retinotopic
organization in human visual cortex and the spatial precision
of functional MRI. Cereb.Cortex 7, 181-192. 1997.
F.Carbonell, L. Galán R. J. Biscay P. Valdés L. Díaz-Comas
K. Worsley M. A. Bobes M. Parra. Detection of event related
potential componets based on the distribution of the maximum
of a T2 random process. Submitted to Clin.Neurophysiol.
Friston, K. J. Frith C. D Turner R. and Franckoviak R.
S. J. Characterizing Evoked Hemodynamics with fMRI. NeuroImage
2, 157-165. 1995a.
Friston, K. J. Frith C. D Franckoviak R. S. J. andTurner
R. Characterizing Evoked Hemodynamics with fMRI: A Multivariate
Approach. NeuroImage 2, 166-172. 1995b.
Friston, K. J. Mechilli R. and Price C. J. Nonlinear Responses
in fMRI: The Balloon Model, Volterra Kernels, and Other
Hemodynamics. NeuroImage 12, 466-477. 2000.
Frostig, R. D. Lieke E. E. Ts'o D. Y. & Ginvald A.
Cortical functional architecture and local coupling between
neuronal activity and the microcirculation revealed by in
vivo high-resolution optical imaging of intrinsic signals.
Proc.Natl.Acad.Sci.USA 87, 6082-6086. 1990.
George, J. S. Aine C. J. Mosher J. C. Schmidt D. M. Ranken
D. M. Schlitt H. A. Wood C. C. Lewine J. D. Sanders J. A.
and Belliveau J. W. Mapping Function in the Human Brain
with Magnetoencephalography, Anatomical Magnetic Resonance
Imaging, and Functional Magnetic Resonance Imaging. Journal
of Clinical Neurophysiology 12(5), 406-431. 1995.
Grinvald, A. Frostig R. D. Siegel R. M. & Bartfeld
E. High resolution optical imaging of functional brain architecture
in the awake monkey. Proc.Natl Acad.Sci.USA 88, 11559-11563.
1991.
Grinvald, A. Lieke E. E. Frostig R. D. and Hildesheim R.
Cortical point-spread function and long-range lateral interactions
revealed by real-time optical imaging of macaque monkey
primary visual cortex. J.Neurosci. 14, 2545-2568.
1994.
Hamalainen, M. S. Hari R. Ilmoniemi R. J. Knuutila J. and
Lounasmaa O. V. Magnetoencephalography-theory, instrumentation,
and applications to noninvasive studies of the working human
brain. Rev.Mod.Physics 65, 413-497. 1993.
Heinze, H. J. Hinrichs H. Scholz M. Burchert W. & Mangun
G. R. Neura; mechanism of global and local processing. A
combined PET and ERP study. J.Cogn.Neurosc. 10, 485-498.
1998.
Heinze, H. J. Mangun G. R. Burchert W. Hinrichs H. Scholtz
M. Munte T. F. Gos A. Scherg M. Johannes S. Hundeshagen
H. et al. Combined spatial and temporal imaging of brain
activity during visual selective attention in humans. Nature
372, 543-546. 1994.
Hoechstetter, K. Rupp A. Meinck H. M. Weckesser D. Bornfleth
H. Stippich C. Berg P. and Scherg M. Magnetic source imaging
of tactile input shows task-independent attention effects
in SII. Cognitive Neuroscience and Neuropsychology 11,
2461-2465. 2000.
Ives, J. R. Warach S. Schmitt F. Edelman R. R. @ Scomer
D. L. Monitoring the patient's EEG during echo-planar functional
MRI in epilepsy. Electroenceph.Clin.Neurophysiol. 87,
417-420. 1993.
Kim, S.-G. Richter W. and Urgubil K. Limitations of Temporal
Resolution in Functional MRI. Magnetic Resonance in Medicine
37, 631-636. 1997.
Korvenoja, A. Huttunen J. Salli E. Pohjonen H. Martinkauppi
S. Palva J. M. Lauronen L. Virtanen J. Ilmoniemi R. J. &
Aronen H. J. Activation of multiple cortical areas in response
to somatosensory stimulation: combined magnetoencephalographic
and functional magnetic resonance imaging. Hum.Brain
Mapp. 8, 13-27. 1999.
Kwong, K. K. Belliveau J. W. Chesler D. A. Goldberg I.
E. Weisskoff R. M. Poncelet B. P. Kennedy D. N. Hoppel B.
E. Cohen M. S. Turner R. et al. Dynamic magnetic resonane
imaging. Proc.Natl.Acad.Sci.USA 89, 5675-5679. 1992.
Liu, A. K. Belliveau J. W. & Dale A. M. Spatiotemporal
imaging of human brain activity using functional MRI constrained
magnetoencephalography data: Monte Carlo simulations. Proc.Natl.Acad.Sci.USA
95, 8945-8950. 1998.
Luck, S. J. Direct and indirect integration of event-related
potentials, functional magnetic resonance, and single-unit
recordings. Hum.Brain Mapp. 8, 115-201. 1999.
MacDonald D, Kabani N, Avis D, Evans AC. Automated 3-D
extraction of inner and outer surfaces of cerebral cortex
from MRI. NeuroImage 2000; 12: 340-356.
Malonek, D. & Grinvald A. Interactions between electrical
activity and cortical microcirculation revealed by imaging
spectroscopy: implications for functional brain mapping.
Science 272, 551-554. 1996.
Malonek, D. Dirnagl U. Lindauer U. Yamada K. Kanno I. &
Grinvald A. Vascular imprints of neuronal activity: relationships
between the dynamics of cortical blood flow, oxygenation,
and volume changes following sensory stimulation. Proc.Natl
Acad.Sci.USA 94, 14826-14831. 1997.
Menon, R. S. Gati J. S. Goodyear B. G. Luknowsky D. C.
& Thomas C. G. Spatial and temporal resolution of functional
magnetic resonance imaging. Biochem.Cell Biol. 76,
560-571. 1998.
Mohamad, D. A. Probabilistic methods for data fusion. Maximum
Entropy Methods. 57-69. 1997. Kluwer Academoc
Publishers.
Ogawa, S. Tank D. W. and Menon R. S. Intrinsic signal changes
accompaning sensory stimulation: functional brain mapping
using MRI. Proc.Natl.Acad.Sci.USA 89, 5951-5955.
1992.
Opitz, B. Mecklinger A. Von Cramon D. Y. & Kruggel
F. Combining electrophysiological and hemodynamic measures
of the auditory odball. Psycophisiology 36, 142-147.
1999.
Pascual-Marqui, R. D. and Michel C. M. LORETA (low resolution
brain electrmagnetic tomography): new authentic 3D functional
imagies of the brain. ISBET Newsl. 1994.
ISBET Newsl.
Richardson, J. M. and Marsh K. A. Basic concepts in multisensor
data fusion. Smith, C. R. et al. eds. Maximum Entropy
Methods. 253-271. 1991. Seatle, Kluwer Academic
Publisher.
Riera J., Fuentes M. E. Valdés P. and Ohárriz Y. EEG Distributed
Inverse Solutions for a Spherical Head Model. Inverse
Problems 14(4), 1009-1019. 1998.
Snyder, A. Z. Abdullaev Y. G. Posner M. I. & Raichle
M. E. Scalp potentials reflect regional cerebral blood flow
responses during processing written words. Proc.Natl
Acad.Sci.USA 92, 1689-1693. 1995.
Stone CJ. Consistent Nonparametric Regression (with discussion).
Annals of Statistics 1977; 5: 595-620.
Valdés-Sosa, P. A. Bosch-Bayard J. B. Aubert-Vázquez E.
F. Virués-Alba T. Morales-Aguilera F. Trujillo-Barreto N.
J. Fuentes-Montero M. E. Soler-McCook J. M. Riera-Díaz J.
J. (CU22550). 10-11-1998. Cuban Patent.
Valdés-Sosa, P. A. García F. Casanova R. Variable resolution
electromagnetic tomography. Aine, C. J. and Okada, Y. Stroink
G. Swithenby S. J. Wood C. C. eds. Biomag 96: Proceedings
of the Tenth International Conference on Biomagnetism.
2000. Springer Verlag.
Warach, S. Ives J. R. Schlaug G. Patel M. R. Darby D. G.
Thangaraj V. Edelman R. R. & Schomer D. L. EEG-triggered
echo-planar functional MRI in epilepsy. Neurology 122,
89-93. 1996.
Worsley KJ. Local maxima and the expected Euler characteristic
of excursion sets of c2 , F and t fields. Advanced Applied
Probability 1994; 26: 13-42.

 Home
Current Issue
Table of Contents
Home
Current Issue
Table of Contents

 indicates that the matrix
indicates that the matrix
 has r rows and c columns.
has r rows and c columns.
 will denote the identity
matrix of order N. For the vector
will denote the identity
matrix of order N. For the vector  ,
,  will denote its transpose.
For a matrix
will denote its transpose.
For a matrix  ,
,  ,
, will denote its determinant and
trace, respectively. The matrix
will denote its determinant and
trace, respectively. The matrix  is the diagonal matrix with
the elements of
is the diagonal matrix with
the elements of  on the main diagonal.
on the main diagonal.  will denote the column vector
formed by stacking all the smaller column vectors of
will denote the column vector
formed by stacking all the smaller column vectors of  . Probability densities will be
expressed according to the notation in Table 3. For example
the r-dimensional multivariate normal density with mean
. Probability densities will be
expressed according to the notation in Table 3. For example
the r-dimensional multivariate normal density with mean
 and covariance matrix
and covariance matrix will be denoted as
will be denoted as  . We shall denote an independent
identically distributed (i.i.d.) random variable
. We shall denote an independent
identically distributed (i.i.d.) random variable  from this distribution is denoted
as
from this distribution is denoted
as  .
.
 is known as the “likelihood”
and
is known as the “likelihood”
and  is the “a priori”
probability density. In Hierarchical Bayesian Modeling the
likelihood and a priori are decomposed into the product
of simpler, known densities. This is the procedure to be
followed below.
is the “a priori”
probability density. In Hierarchical Bayesian Modeling the
likelihood and a priori are decomposed into the product
of simpler, known densities. This is the procedure to be
followed below.

 EEG electrodes and
EEG electrodes and
 MEG sensors respectively
for each time instant
MEG sensors respectively
for each time instant
 ;
; and
and
 are the vectors of measurement
errors for the EEG and MEG sensor respectively.
are the vectors of measurement
errors for the EEG and MEG sensor respectively.
 ,
,
 are the electric and magnetic
lead field matrices multiplied by the known vectors of orientations
of the current density for each point in the gray matter.
In other words it has been assumed that the orientation
of the current density vector for each possible source is
constant in time and directed perpendicular to the cortex.
Thus, only the strength and sign of neuronal PCD
are the electric and magnetic
lead field matrices multiplied by the known vectors of orientations
of the current density for each point in the gray matter.
In other words it has been assumed that the orientation
of the current density vector for each possible source is
constant in time and directed perpendicular to the cortex.
Thus, only the strength and sign of neuronal PCD  points in the solution grid
will be estimated. A further assumption is that,
points in the solution grid
will be estimated. A further assumption is that, i.i.d
i.i.d
 ,
,
 are the sensor noise variances.
From these expressions it follows that the likelihoods for
the electrophysiological measurements read:
are the sensor noise variances.
From these expressions it follows that the likelihoods for
the electrophysiological measurements read:
 , is convolved with a known hemodynamic
matrix response function
, is convolved with a known hemodynamic
matrix response function  [Friston 1995a;Friston 1995b]:
[Friston 1995a;Friston 1995b]:


 .
. ,
is a sub-sampled version of
,
is a sub-sampled version of
 (since the acquisition rate
of the MRI scanner is much lower than that of electrophysiological
measurements). This is formalized by use of the sub-sampling
matrix
(since the acquisition rate
of the MRI scanner is much lower than that of electrophysiological
measurements). This is formalized by use of the sub-sampling
matrix 

 is the number of fMRI
time samples. We shall assume that the fMRI measurement
errors are distributed as
is the number of fMRI
time samples. We shall assume that the fMRI measurement
errors are distributed as i.i.d.
i.i.d.
 .
We shall assume that both the
PCD and the NMD at each voxel are generated by some deterministic
or stochastic mechanisms
.
We shall assume that both the
PCD and the NMD at each voxel are generated by some deterministic
or stochastic mechanisms  and
and  such that:
such that:

 as well as for all the hyperparameters
by means of the densities
as well as for all the hyperparameters
by means of the densities  ,
,
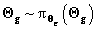 and
and
 . Estimation of all these parameters
may be carried out by any of the usual methods of Bayesian
computation.
. Estimation of all these parameters
may be carried out by any of the usual methods of Bayesian
computation. and
and
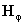 as well as
as well as
 , then the specific formulation
of model could be relatively straightforward. Some advances
have been made in terms of formulating the pair of equations
(in our notation)
, then the specific formulation
of model could be relatively straightforward. Some advances
have been made in terms of formulating the pair of equations
(in our notation)  and
and
 [Friston 2000]. While of
great promise this line of modeling is not yet complete
for practical application.
[Friston 2000]. While of
great promise this line of modeling is not yet complete
for practical application.
 that is equivalent to assuming that:
that is equivalent to assuming that:

 a constant introduced to
match the units of PCD and NMD,
a constant introduced to
match the units of PCD and NMD,
 and
and
 denoting i.i.d.
denoting i.i.d.
 variables and
variables and
 the Hadamaard product.
As a consequence of this specification the a priori densities
of the PCD and NMD are:
the Hadamaard product.
As a consequence of this specification the a priori densities
of the PCD and NMD are:


 is taken as an Inverse Wishart:
is taken as an Inverse Wishart:
 is a scaling factor, and
is a scaling factor, and
 is are the degrees of freedom
In order to completely specify our model, we include non-informative
inverse Gamma distributions for the hyperparameters:
is are the degrees of freedom
In order to completely specify our model, we include non-informative
inverse Gamma distributions for the hyperparameters: ,
,
 ,
,
 ,
,
 ,
,


 that maximize . These are
found by the Iterated Conditional Maximization method for
finding the Maximum A Posteriori (MAP) estimators for the
parameters. We iterate between the following expressions:
that maximize . These are
found by the Iterated Conditional Maximization method for
finding the Maximum A Posteriori (MAP) estimators for the
parameters. We iterate between the following expressions: