Linear inverse estimation
of cortical sources by using high resolution EEG and
fMRI priors
Babiloni, F.a,
Babiloni, C.ad, Carducci, F.ad,
Angelone, L.a, Del Gratta, C.bf,
Romani, GL.bf, Rossini, PM.c, and
Cincotti, F.ae
Corresponding author: Dr.
Fabio Babiloni, Dipartimento di Fisiologia Umana e Farmacologia,
Università di Roma “La Sapienza” P.le A. Moro
5, 00185 Roma, Italy
Email: fabio.babiloni@uniroma1.it, Tel: +39 06 4991 0317 Fax: +39 06 4991
0917
1. Introduction
It is well known that conventional EEG (i.e.
19 scalp electrodes disposed according to 10-20 International
system) offers a high temporal resolution (milliseconds),
but exhibits a low spatial resolution (about 6-9 cm) as
an image of the cortical sources generating the potential.
This is due to the different conductivities of cortex, dura
mater, skull, and scalp (Gevins 1989; Nunez, 1995). The
distortion of the recorded scalp potential distribution
is further increased by the ears and eyeholes, which represent
shunt paths for intra-cranial currents (Nunez, 1981, 1995).
Another factor that affects the scalp potentials is the
position and dynamics of electrical reference used during
EEG recordings, which may filter out spatial components
of the potential distribution over the scalp (Nunez, 1981).
On the whole, the scalp potential distribution is a “blurred”
copy of the original cortical potential distribution. Remarkably,
the increase of spatial sampling (to 64 or 128) is not sufficient
per se to enhance the spatial information content
of scalp potential distribution (Gevins et al., 1999; Nunez,
1995).
Neural sources of EEG can be localized by
making a priori hypothesis on their number and extension.
When the EEG activity is mainly generated by a known number
of cortical sources (i.e. short-latency evoked potentials),
the location and strength of these sources can be reliably
estimated by the dipole localization technique (Scherg and
von Cramon, 1984). However, except for the processing of
sensory inputs from the primary sensory cortical areas,
the activity of multiple extended cortical sources has been
reported in a variety of human tasks, i.e. simple movement
planning and execution as well as complex memory tasks (Carlson
et al., 1998). In these cases, the source activity could
be estimated by using extended source models and spherical
or realistic head volume conductor models in the context
of linear inverse theory (Dale and Sereno, 1993; Pascual-Marqui
et al. 1994; Grave de Peralta and Gonzales Andino 1998;
Babiloni et al., 2000a; Dale et al., 2000). With this approach,
thousands of equivalent current dipoles are used as a source
model and realistic head models reconstructed from magnetic
resonance images (MRIs) serve as a volume conductor (Meijs
et al., 1993; Yvert et al. 1995). Regularized linear inverse
solution attributes a strength value at each dipole of the
source model from the scalp potential distribution.
The solution space (i.e. the set of all possible
combinations of the cortical dipoles’ strengths) is
generally reduced by using geometrical constraints, for
instance by using cortical dipoles oriented according to
the normal direction of the cortical surface. (Dale and
Sereno, 1993). Constraints on the energy of the estimated
cortical activity are decisive to reduce further the solution
space (minimum-norm solution; Hamalainen and Ilmoniemi,
1984, Dale and Sereno, 1993). Recently, the solution space
has been restricted by using information deriving from cerebral
blood flow measures (Liu et al., 1998; Ahlfors et al., 1999;
Babiloni et al., 2000b; Dale et al., 2000; Liu, 2000).
The rationale of such multimodal approach is that neural
activity generating EEG potentials increases glucose and
oxygen demands (Magistretti et al., 1999). This results
in an increase in the local hemodynamic response that can
be modeled by functional magnetic resonance images (fMRI;
Grinvald et al., 1986; Puce et al., 1997). On the whole,
such a correlation between electrical and hemodynamical
concomitants provides the basis for a spatial correspondence
between fMRI responses and EEG source activity.
In this paper, we present two methods for
the modeling of human cortical activity from combined high-resolution
electroencephalography (EEG) and functional magnetic resonance
imaging (fMRI) data. These methods were based on linear
inverse estimation and included subject’s multi-compartment
head model (scalp, skull, dura mater, cortex) constructed
from magnetic resonance images and a multi-dipole source
model. Information on the hemodynamic responses of the investigated
cortical areas derived from block-design and event-related
functional Magnetic Resonance Imaging (fMRI) were used as
a priors in the resolution of the linear inverse problem
used for the estimation of the cortical activity. High resolution
EEG (128 electrodes) and fMRI data, were recorded in separate
sessions, while normal subjects executed voluntary right
one-digit movements.
2. Methods
Realistic head and source models
Sixty-four T1-weighted sagittal Magnetic Resonance
(MR) images were acquired (30 ms repetition time, 5 ms echo
time, and 3 mm slice thickness without gap) of the experimental
subjects’ head. These images were processed with 3D
segmentation and triangulation algorithms for the construction
of a model reproducing scalp, skull, and dura mater surfaces
with about 1000 triangles for each surface. Source models
were built with the following procedure: (i) the voxels
belonging to the MR volume of the cortex were selected with
a semiautomatic procedure (threshold algorithm); (ii) these
points were triangulated obtaining a fine mesh with about
100,000 triangles; (iii) a coarser mesh was obtained by
resampling the one described above down to about 6,000 triangles,
taking care that the general features of the neocortical
envelope were well preserved especially in correspondence
of pre- and post-central gyri and frontal mesial area; (iv)
an orthogonal unitary equivalent current dipole was placed
in each node of the triangulated surface, with direction
parallel to the vector sum of the normals to the surrounding
triangles.
EEG linear inverse estimation
Taking into account the measurement noise
n, supposed to be normally distributed, an estimate
of the dipole source configuration that generated a measured
potential b can be obtained by solving the linear
system:
 (1)
(1)
where A is a matrix with number of
rows equal to the number of sensors and number of columns
equal to the number of modeled sources, called lead field
matrix. The electrical lead field matrix A and the
data vector b must be referenced consistently. Among
the several equivalent solutions for the underdetermined
system (1), the solution
was chosen that satisfies the following variational problem
for the sources x (Dale and Sereno, 1993; Grave de
Peralta and Gonzalez Andino, 1998; Liu, 2000):
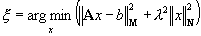 (2)
(2)
where M, N are the matrices
associated to the metrics of the data and of the source
space, respectively. The solution of the variational problem
depends on adequacy of the data and source space metrics.
Under the hypothesis of M and N positive definite,
the solution of eq. 2
is given by computing the pseudoinverse matrix G
according to the following expressions:
 ,
,
 (3)
(3)
An optimal regularization of this linear system
was obtained by the L-curve approach (Hansen, 1992). This
curve, which plots the residual norm versus the solution
norm at different l values, was used to choose the optimal
amount of regularization in the solution of the linear inverse
problem. Computation of the L-curves and optimal l correction values was performed with the original Hansen’s
routines (Hansen, 1994). The metric M, characterizing
the idea of closeness in the data space, can be particularized
by taking into account the sensors noise level by using
the Mahalanobis distance (Grave de Peralta and Gonzalez
Andino, 1998). The source metric N can be particularized
by taking into account the information from the hemodynamic
responses of the single voxels, as showed in the following
section.
Functional hemodynamical
coupling and linear inverse estimation of source activity
Here, we present two characterizations of
the source metric N that can provide the basis for
the inclusion of the information about the statistical hemodynamic
activation of i-th cortical voxels into the linear
inverse estimation of the cortical source activity. The
first characterization of the source metric N takes
into account all the cortical voxels on the basis of their
electrical “closeness” to the EEG sensors (column
norm normalization; Pascual-Marqui et al., 1994; Gorozdnistky
et al., 1995; Grave de Peralta and Gonzalez Andino, 1998).
In this case, the inverse of the resulting source metric
is
 (4)
(4)
in which  is the i-th element of the inverse
of the diagonal matrix N and
is the i-th element of the inverse
of the diagonal matrix N and  is the L2 norm of the i-th column
of the lead field matrix A.
is the L2 norm of the i-th column
of the lead field matrix A.
Introducing fMRI priors into the linear inverse
estimation produces a bias in the solution: statistically
significantly activated fMRI voxels, which are returned
by the so called percentage change approach (Kim et al.,
1993), are weighted to account for the EEG measured potentials.
The inverse of the resulting metric is
 (5)
(5)
in which  and
and  has the same meaning described above.
The
has the same meaning described above.
The  is
a function of the statistically significant percentage increase
of the fMRI signal assigned to the i-th dipole of
the modeled source space. This function was expressed as
is
a function of the statistically significant percentage increase
of the fMRI signal assigned to the i-th dipole of
the modeled source space. This function was expressed as
 ,
,
 ,
,  (6)
(6)
where  is the percentage increase of the fMRI
signal during the task state for the i-th voxel and
the factor K tunes fMRI constraints in the source
space. Fixing K = 1 let us disregard fMRI
priors, thus returning to a purely electrical solution;
a value for K >> 1 allows only the
sources associated with fMRI active voxels to participate
in the solution. It was shown that a value for K
in the order of 10 (90% of constraints for the fMRI information)
is useful to avoid mislocalization due to over constrained
solutions (Liu et al., 1998; Dale et al., 2000; Liu, 2000).
In the following the estimation of the cortical activity
obtained with this metric will be denoted as diag-fMRI.
is the percentage increase of the fMRI
signal during the task state for the i-th voxel and
the factor K tunes fMRI constraints in the source
space. Fixing K = 1 let us disregard fMRI
priors, thus returning to a purely electrical solution;
a value for K >> 1 allows only the
sources associated with fMRI active voxels to participate
in the solution. It was shown that a value for K
in the order of 10 (90% of constraints for the fMRI information)
is useful to avoid mislocalization due to over constrained
solutions (Liu et al., 1998; Dale et al., 2000; Liu, 2000).
In the following the estimation of the cortical activity
obtained with this metric will be denoted as diag-fMRI.
The previous definition of the source metric
N results in a matrix in which the off-diagonal elements
are zero (diag-fMRI). However, we can take advantage of
the off-diagonal elements of the matrix N to insert
the information about the functional coupling of the cortical
sources. In particular we set the generic ij entry
of the inverse of matrix N as in the following
 (7)
(7)
where  and
and  have the same meaning described above
and
have the same meaning described above
and  is
the degree of functional coupling between the source i
and the source j during the particular task analyzed.
Such coupling was revealed by the correlation of their hemodynamic
responses obtained by the event-related fMRI data. In the
following the estimation of the cortical activity obtained
with this metric will be denoted as corr-fMRI. It is of
interest that in the case of uncorrelated sources (
is
the degree of functional coupling between the source i
and the source j during the particular task analyzed.
Such coupling was revealed by the correlation of their hemodynamic
responses obtained by the event-related fMRI data. In the
following the estimation of the cortical activity obtained
with this metric will be denoted as corr-fMRI. It is of
interest that in the case of uncorrelated sources ( ,
,  ;
;  ), the corr-fMRI
formulation leads back to the diag-fMRI one. Fig.1 summarizes
the different approaches pursued here in order to insert
the hemodynamical constraints in the solution of the linear
inverse problem for the estimation of the cortical sources
of the recorded EEG in a unique mathematical formulation.
), the corr-fMRI
formulation leads back to the diag-fMRI one. Fig.1 summarizes
the different approaches pursued here in order to insert
the hemodynamical constraints in the solution of the linear
inverse problem for the estimation of the cortical sources
of the recorded EEG in a unique mathematical formulation.

Figure 1. Upper part: Estimate of the hemodynamic
coupling between two generic cortical sources (i-th and
j-th) as obtained by the computation of the cross-correlation
between the waveforms of the fMRI responses. These waveforms
(Si, Sj) were obtained during a simple voluntary movement
(right middle finger extension). Lower part: mathematical
formulation of the inverse of the source metric N to be
used in the solution of the linear inverse problem. Corr(Si,Sj)
is the zero-lag correlation between the two hemodynamic
waveforms Si and Sj, and is the Kroneker symbol.
EEG recording
EEG activity was recorded (0.1-100 Hz bandpass)
with 128 electrodes (linked earlobe electrical reference).
Electrode positions and reference landmarks were digitized
for subsequent integration between the EEG, MEG, and MR
data. Electrooculogram (EOG; 0.1-100 Hz bandpass) and electromyogram
(EMG; 1-100 Hz bandpass) from m. extensor digitorum
of both sides were also recorded. EOG served to control
blinking/eye movements and EMG to control operating muscle
response and involuntary mirror movements. All data were
acquired (400 Hz sampling rate) from 3 sec before to 1 sec
after the onset (zerotime) of the EMG response from the
operating muscle. About 200 single trials were collected
and averaged for each subject.
3. Results and Discussion
Fig. 2 illustrates
the topographic map of readiness potential distribution
recorded at the scalp about 200 ms before a right middle
finger extension (subject #1). Note the extension of the
maximum of the negative scalp potential distribution, roughly
overlying frontal and centroparietal areas contralateral
to the movement. Fig.2 illustrates also the percent values
of the fMRI response during the movement in a separate experimental
session. The maximum values of the fMRI responses are located
in the voxels roughly corresponding to the primary somatosensory
and motor areas (hand representation) contralateral to the
movement.

Figure 2. Left: scalp
potential distribution recorded about 200 msec before the
movement onset (128 recording channels) in a separate session.
This distribution is representative of the so called readiness
potential. Percent color scale in which maximum negativity
(-100%) is coded in red and maximum positivity (+100%) is
coded in black.. Right: fMRI response related to the movement
in subject 1. Only the brain voxels whose hemodynamic response
is increased statistically are shown. The fMRI response
is integrated to a MRI-based reconstruction of the cortical
surface. The red to yellow color bar codes the percentage
of the increase of the fMRI response
Fig. 3 illustrates
the cortical distribution of the current density
estimated with linear inverse approach from the potential
distribution of Fig.2. Such an approach used no-fMRI constraint
as well as two types of fMRI constraints, i.e. one based
on block-design (diag-fMRI) and the other on event-related
design (corr-fMRI). The cortical distributions are represented
on the realistic subject’s head volume conductor model.
Linear inverse solutions obtained with the fMRI priors (diag
and corr-fMRI) present more localized spots of activations
with respect to those obtained with the no fMRI priors.
Remarkably, the spots of activation were localized in the
hand region of the primary somatosensory (post-central)
and motor (pre-central) areas contralateral to the movement.
In addition, spots of minor activation were observed in
the frontocentral medial areas (including supplementary
motor area) and in the primary somatosensory and motor areas
of the ipsilateral hemisphere. Similar results were
obtained in the other main components of the movement-related
potentials (i.e. motor potentials and movement-evoked potential)
and in the other subject.

Figure 3. Cortical distributions
of the current density estimated with a linear inverse approach
from the readiness potential of Fig.2. Linear inverse estimates
are obtained with no fMRI constraints (no-fMRI) and two
kinds of fMRI constraints, one based on the strengths of
the cortical fMRI responses (diag-fMRI) and the other on
the correlation between fMRI responsive cortical areas (corr-fMRI).
Percent color scale: maximum negativity (-100%) is coded
in red and maximum positivity (+100%) is coded in black.
The results of the present study are in line
with those regarding the coupling between cortical electrical
activity and hemodynamic measure as indicated by a direct
comparison of maps obtained using voltage-sensitive dyes,
which reflect depolarization of neuronal membranes in superficial
cortical layers, and maps derived from intrinsic optical
signals, which reflect changes in light absorption due to
changes in blood volume and oxygen consumption (Shoham et
al., 1999). Furthermore, previous studies on animals have
also shown a strong correlation between local field potentials,
spiking activity, and voltage-sensitive dye signals (Arieli
et al., 1996). Finally, studies in humans comparing the
localization of functional activity by invasive electrical
recordings and fMRI have provided evidence of a correlation
between the local electrophysiological and hemodynamic responses
(Puce et al, 1997). This may suggest that the local fMRI
responses can be reliably used to bias the estimation of
the electrical activity in the regions showing a prominent
hemodynamic response.
It may be argued that combined EEG-fMRI responses
could be less reliable for the modeling of cortical activation
in the case of a spatial mismatch between electrical and
hemodynamical responses. However, previous studies have
suggested that by using the fMRI data as a partial constraint
in the liner inverse procedure, it is possible to obtain
accurate source estimates of electrical activity even in
the presence of some spatial mismatch between the generators
of EEG data and the fMRI signals (Liu et al., 1998; Liu,
2000). Furthermore, it is questionable if the level
of the bias for the hemodynamical constraints in the linear
inverse estimation can be the same with the diag-fMRI and
corr-fMRI approaches, (so did we in this present study).
We think that this issue deserves a specific simulation
study, using the literature indexes capable of assessing
the quality of the linear inverse solutions (Pascual Marqui
et al., 1994; Grave de Peralta and Gonzalez Andino, 1998,
Babiloni et al., 2001).
In conclusion, the present paper dealt with
the issue of combining fMRI and EEG data for the study of
event-related cortical responses. This approach can be further
enriched incorporating magnetoencephalographic data in the
linear inverse estimation, to constitute an unsurpassable
non invasive technology for the analysis of human higher
brain functions at a high temporal and a good spatial resolution.
References
Ahlfors SP, Simpson GV, Dale AM, Belliveau
JW, Liu AK, Korvenoja A, Virtanen J, Huotilainen M, Tootell
RB, Aronen HJ, Ilmoniemi RJ (1999). “Spatiotemporal
activity of a cortical network for processing visual motion
revealed by MEG and fMRI”. J Neurophysiol, 1999
Nov, 82(5): 2545-55.
Arieli A, Sterkin A, Grinvald A, Aertsen
AD (1996). “Dynamics of Ongoing Activity: Explanation
of the Large Variability in Evoked Cortical Responses”,
Science 273: 1868‑71.
Babiloni F., Babiloni C., Locche L., Cincotti F., Rossini
P.M., Carducci F. (2000a) "High-resolution electro-encephalogram:
source estimates of Laplacian-transformed somatosensory-evoked
potentials using a realistic subject head model constructed
from magnetic resonance images." Med Biol Eng Comput.
Sep;38(5):512-9.
Babiloni F, Carducci F, Cincotti F, Del
Gratta C, Roberti GM, Romani GL, Rossini PM, Babiloni C
(2000b). “Integration of High Resolution EEG and Functional
Magnetic Resonance in the Study of Human Movement-Related
Potentials”, Method Inform Med 39: 179–82.
Babiloni F., Babiloni C., Carducci F., Angelone C.,,
Del Gratta C., Romani G.L, Rossini, P.M. and Cincotti
F. “Multimodal integration of high resolution EEG and
functional Magnetic Resonance: a simulation study”;
Human Brain Mapping Conference, Brighton, 12-16 June
2001.
Carlson, S.,
Martinkauppi, S., Rama, P., Salli, E., Korvenoia, A. and
Aronen, H. (1998) "Distribution of cortical activation
during visuospatial n-back tasks as revealed by functional
magnetic resonance imaging." Cerebral Cortex,
8:743-752
Dale A, Liu A, Fischl B, Buckner R, Belliveau
JW, Lewine J, Halgren E (2000). “Dynamic Statistical
Parametric Mapping: Combining fMRI and MEG for High-Resolution
Imaging of Cortical Activity”, Neuron, 26: 55‑67.
Dale AM and Sereno M (1993). “Improved
localization of cortical activity by combining EEG and MEG
with MRI cortical surface reconstruction: a linear approach”,
J. Cognitive Neuroscience, 5: 162‑76.
Dale A, Liu A, Fischl B, Buckner R, Belliveau
JW, Lewine J, Halgren E (2000). “Dynamic Statistical
Parametric Mapping: Combining fMRI and MEG for High-Resolution
Imaging of Cortical Activity”, Neuron, 26: 55‑67.
Gevins, A. (1989) "Dynamic functional topography of
cognitive task". Brain Topogr., 2: 37-56.
Gevins A, Le J, Leong H, McEvoy LK, Smith ME (1999) "Deblurring",
J Clin Neurophysiol 16(3):204-13
Gorodnitsky IF, George JS and Rao BD (1995).
“Neuromagnetic source imaging with FOCUSS: a recursive
weighted minimum norm algorithm”. Electroencephalogr
Clin Neurophysiol. 95: 231-51.
Grave de Peralta Menendez R and Gonzalez
Andino SL (1998). “A critical analysis of linear inverse
solutions to the neuroelectromagnetic inverse problem”,
IEEE Trans Biomed Eng, 45(4):440-8
Grave de Peralta R and Gonzalez Andino SL
(1998). “Distributed source models: standard solutions
and new developments”. In: Uhl, C. (ed): Analysis
of neurophysiological brain functioning. Springer Verlag,
pp.176‑201.
Grinvald A, Lieke E, Frostig RD, Gilbert
CD and Wiesel, TN (1986). “Functional architecture
of cortex revealed by optical imaging of intrinsic signals”.
Nature 324: 361‑4.
Hämäläinen M and Sarvas J (1989). “Realistic
conductivity geometry model of the human head for interpretation
of neuromagnetic data”. IEEE Trans Biomed Eng
36: 165‑71.
Hämäläinen, M. and Ilmoniemi,
R. (1984) "Interpreting meausured magnetic field of
the brain: estimates of the current distributions".
Technical report TKK-F-A559, Helsinki University
of Technology
Hansen PC (1992). “Analysis of discrete
ill-posed problems by means of the L-curve”. SIAM
Review 34: 561‑80.
Hansen PC (1994). “Regularization tools,
a Matlab package for analysis and solution of discrete ill-posed
problems”. Numer Algorithms 6: 1‑35.
Kim SG, Ashe J, Georgopoulos AP, Merkle
H, Ellermann JM, Menon RS, Ogawa S, Ugurbil K (1993). “Functional
imaging of human motor cortex at high magnetic field”,
J Neurophysiol, 69(1): 297-302.
Liu AK (2000). “Spatiotemporal brain
imaging”. PhD dissertation, Massachusetts Institute
of Technology, Cambridge, Massachusetts.
Liu AK, Belliveau JW and Dale AM (1998).
“Spatiotemporal imaging of human brain activity using
functional MRI constrained magnetoencephalography data:
Monte Carlo simulations”, Proc Natl Acad Sci
USA, 95(15): 8945‑50.
Magistretti PJ; Pellerin L; Rothman DL;
Shulman RG (1999). “Energy on demand”, Science,
283(5401): 496-7
Meijs J, Weier O, Peters M, and van Oosterom
A (1993). “On the numerical accuracy of the boundary
element method”, IEEE Trans. Biomed. Eng. 42:
1038-1049
Nunez P (1981). “Electric fields of
the brain”. New York :Oxford University Press.
Nunez P (1995). “Neocortical dynamics
and human EEG rhythms”. New York: Oxford University
Press. 722p.
Pascual Marqui RD, Michel CM and Lehmann
D (1994). “Low resolution electromagnetic tomography:
a new method for localizing electrical activity of the brain”
Int. Journ. of Psychopsysiology, 18: 49‑65.
Puce A, Allison T, Spencer SS, Spencer DD
and McCarthy G (1997). “Comparison of cortical activation
evoked by faces measured by intracranial field potentials
and functional MRI: two case studies”. Hum Brain
Mapp, 5: 298‑305.
Scherg M, von Cramon D and Elton M (1984).
“Brain-stem auditory-evoked potentials in post-comatose
patients after severe closed head trauma”, J Neurol,
231(1): 1-5
Shoham D, Glaser DE, Arieli A, Kenet T,
Wijnbergen C, Toledo Y, Hildesheim R and Grinvald A. (1999).
“Imaging cortical dynamics at high spatial and temporal
resolution with novel blue voltage-sensitive dyes”.
Neuron 24: 791–802
Yvert B, Bertrand O, Echallier JF and Pernier
J (1995). “Improved forward EEG calculations using
local mesh refinement of realistic head geometries”,
Electroencephalogr Clin Neurophysiol, 95(5): 381-92

 Home
Current Issue
Table of Contents
Home
Current Issue
Table of Contents
