O. Blanc*, N. Virag*, O. Egger*, J.-M. Vesin*, and L. Kappenberger**
* Signal Processing Laboratory, Swiss Federal Institute of Technology, Lausanne, Switzerland
** Division of Cardiology, State University Hospital, Lausanne, Switzerland
Introduction
Computer models of electrical propagation in
the heart represent a powerful tool to understand the mechanism of arrhythmias.
Their major drawback is their heavy computational load, which imposes restrictions
for realistic modeling. Models are designed as tradeoff between membrane
complexity, tissue structure, tissue size and duration of simulation. The
most effective computer models of the heart have up to 1'000'000 spatial
nodes, thus heavily restraining the size of tissue that can be simulated
in 2D or 3D. Stronger restrictions occur when the objective is the realistic
simulation of arrhythmia, because of the important amount of cardiac cells
involved (the whole ventricular or atrial myocardium), and the several
seconds of time required to reach a fully developed arrhythmic state. Models
of conduction in the cardiac tissue based on membrane ionic currents have
been developed over the last years, mostly for two dimensional tissue [1],
but very few models take into account the geometry of the heart, which
is a key factor for the development of arrhythmias. To overcome this problem,
models incorporating anatomical 3D properties mainly use simplified membrane
kinetics, such as the FitzHugh-Nagumo model [2].
While in the ventricles, reentry is known to be a 3D process involving
transmural activity, major atrial activations involve only the thin walls
of the atria and therefore the arrhythmic process could be simplified to
a 2D problem in a 3D structure [2].
Based on this assumption, we describe in this paper a computer model
of the human atria, using a simplified 3D geometry with a realistic atrial
topology and size and a physiological cellular model (Beeler-Reuter or
Luo-Rudy). The objective of this paper is to show how this anatomical computer
model of human atria can contribute to the understanding of atrial arrhythmias
and provide a tool for the development of therapeutic strategies.
Methods
Cardiac Tissue Model
The cardiac tissue consists of a grid of cardiac cells interconnected
via gap junctions. Electrical propagation in this tissue can be modeled
using a cable equation. If we neglect current flow in the extracellular
domain, we obtain the monodomain formulation of action potential propagation,
which is described by the following reaction-diffusion equation:
| |
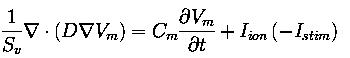 |
(1) |
where Sv is the cell membrane surface to cell volume
ration, D the conductivity tensor, Vm the transmembrane
potential, Cm the membrane capacitance, Iion
the sum of the transmembrane ionic currents and Istim
a stimulus current. All simulations where conducted with Cm =
1 µF/cm2 and Sv = 0.24 µm-1.
Intracellular resistivities can be individually programmed in the longitudinal
and transversal directions via the conductivity tensor D (D
is assumed to be diagonal), allowing introduction of heterogeneities in
the tissue. In the presented results, the cardiac tissue is assumed to
be globally homogeneous and isotropic. Heterogeneities have been introduced
only to model major conductive obstacles like veins and valves or local
slow conduction regions (between tricuspid valve and inferior vena cava
for example). The sum of the transmembrane ionic currents Iion
in
Equation (1) is computed either with the Beeler-Reuter
[3]
or the Luo-Rudy [4] models. These models are described
by non-linear equations involving voltage dependent closing and opening
gating variables for the different ion channels. In this paper we present
simulations obtained only with the Beeler-Reuter model but similar results
have been obtained with the Luo-Rudy formulation for Iion.
Equation (1) has been discretized by
a finite differences method in both time and space, and solved in two steps:
firstly an explicit computation of the membrane ionic currents Iion,
using Runge-Kutta integration and lookup tables to speed up gating variable
computation, and secondly a semi-implicit computation of the remaining
diffusive terms through a classical ADI scheme. The semi-implicit computation
has been preferred to a simpler fully explicit method due to its stability
even if a comparison between ADI and explicit methods show that it introduces
a computational load increase of 30% approximately.
The choice of time and space discretizations of Equation
(1) is the result of a tradeoff between computation speed and accuracy.
Computer simulations of electrical propagation and especially reentry simulations
impose strong requirements on the spatial discretization, which in turn
has a major impact on the computational requirements. Typical signatures
of incorrect spatial discretization are [5]: reduced
propagation velocity (sometimes leading to artificial block of propagation),
artificial anisotropy depending on the grid directions with slower propagation
along the grid directions than in the diagonal directions (square waves
patterns), and alteration of the reentry dynamics. The two main restrictions
to spatial refinements are computation time and memory requirements (and
numerical stability if explicit solving schemes were used). Based on these
considerations, we have used space steps of 200 µm and fixed time
steps of 25 µs. The resistivities (inverse of conductivity) have
been varied between 80 Ohm·cm to 800 Ohm·cm, corresponding
to propagation velocities ranging from approximately c = 90 cm/s
to 30 cm/s. From a computational point of view, a reduction of propagation
velocity by a resitivity increase is equivalent to a dilation of both atria
while keeping a normal propagation velocity.
Several experiments have been performed to show the realistic behavior
of this two-dimensional tissue [6].
Human Atria Model
Based on the assumption that atria are constituted of thin walls [2],
we have built our anatomic model of human right and left atrium by folding
the described two-dimensional cardiac tissue model into a 3D structure
of one layer of cells. The human atria are modeled by a simplified structure
composed of two connected ellipsoids with a diameter of 4.5 cm and an overall
length of 8 cm. Non conductive obstacles are placed to simulate the veins
and the valves, as shown in Figure 1. The total
area of the structure is approximately 100 cm2, with the orifices
accounting for 20% of this area. The surface of the mitral (MV) and tricuspid
(TV) valves is 4 cm2, 2 cm2 for the superior vena
cava (SVC), 2.5 cm2 for the inferior vena cava (IVC) and 1 cm2
for each pulmonary vein (PV). These values are comparable with those found
in human atria.
In spite of its crude approximation, our atrial geometry presents all
major anatomical obstacles important for the simulation of atrial arrhythmias.
Furthermore, it has a realistic size with a reasonable spatial discretization.
Further work will enable us to develop a more accurate anatomical structure
and geometry of the atria including pectinate muscles and a septal structure.

|
| (a) | (b) |
Figure 1. Geometry of the proposed human atria model with the holes
represented in black, (a) anterior view with Superior Vena Cava (SVC) and
Sino-Atrial Node (SA Node) visible in the right atrium, (b) posterior view
with Inferior Vena Cava (IVC) and Tricuspid Valve (TV) visible in the right
atrium, Pulmonary Veins (PV) and Mitral Valve (MV) visible in the left
atrium.
Electrical activation is initiated from selected regions by injecting
an intracellular current (Istim in Equation
(1)) to simulate periodic depolarization of the sino-atrial (SA) node,
or premature stimuli. Furthermore, different action potential modifications
have been tested by modulation of the ionic currents, simulating membrane
defects or electrical disturbances.
The model has been implemented in C++ with double precision and runs
on multiple platforms, including Windows PC's, cluster of Linux PC's, and
a SGI Cray Origin2000 with 38 64-bits processors. The computation of 1
ms of real time typically requires 1 min of CPU with a single Pentium II
processor.
Results
Sinus Rhythm Propagation
Normal propagation was initiated from the SA node region. From there
the activation wavefront propagates over the right to the left atrium.
In the model of healthy tissue, the propagation of a normal sinus beat
results in a total activation time for the atria of about 90 ms. The propagation
speed is 90 cm/s and the action potential duration is 289 ms at a cycle
length of 1000 ms. This propagation velocity is in the range of values
reported for physiological tissues
[7]. A simulation
of 5 sinus rhythm beats at a rate of 60 bpm is presented real time in
Video
1.
Video 1: Sinus rhythm propagation at 60 bpm in a healthy atrial
tissue (propagation velocity c = 90 cm/s). The video is displayed real
time and the total depolarization time for both atria is 90 ms.
(The VLC Media Player  is recommended for viewing the video.)
is recommended for viewing the video.)
Atrial Arrhythmias
Several experiments of arrhythmia initiation have been performed, using
a programmed stimulation protocol as in electrophysiological studies. S1
was the basic drive impulse initiated from the SA node at a cycle length
of 1000 ms. Ectopic beats S2 and S3 (area 1.5 mm2,
intensity equal to the double of the fully repolarized threshold) have
been introduced with various coupling intervals and at several locations.
In the healthy tissue, defined with a normal anatomy and size, all initiation
attempts have failed since no sustained arrhythmia could be seen. Sustained
reentry could not exist in the healthy tissue model due to wavelength constraints:
an action potential with a duration of 289 ms and a propagation velocity
of 90 cm/s leads to a wavelength of 27 cm, which is much larger than the
atrial dimensions. Therefore, at basic conditions, this model seems electrically
stable. Arrhythmias could be initiated when the wavelength was decreased
to a value lower than the atria dimensions. First, the propagation velocity
was reduced from 90 cm/s to 30 cm/s, either globally in the whole tissue,
or locally (to mimic slow conduction in the isthmus between IVC and TV
for example) by increasing the resistivity from 80 Ohm·cm to 800
Ohm·cm. As already mentioned, this decreased propagation velocity
can also be interpreted as a normal propagation velocity in dilated atria.
Furthermore, a decrease of action potential duration by an inhibition of
the Is current in the Beeler-Reuter model facilitates
reentry because of an action potential shortening.
Even in a tissue with decreased wavelength, most programmed stimulation
protocols have led to non sustained arrhythmias. Only critically timed
and located premature beats could initiate atrial flutter or atrial fibrillation.
Atrial flutter has been induced by applying one single ectopic beat S2
between IVC and TV carefully timed after a sinus rhythm propagation. This
leads to a single periodic reentrant wavefront anatomically anchored around
IVC and TV. These results are similar to those reported in [2].
Atypical flutter can also be induced with premature beats in the PV region
by using the same technique. The average rate of flutter in our model is
250 bpm and the periodic structure is clearly visible.
The initiation of atrial fibrillation in the model is more complex
than flutter initiation. Several (two or more) carefully located and timed
ectopic beats following a normal sinus rhythm depolarization are required
to reach a fibrillatory state. Furthermore fibrillation could only be initiated
in a tissue with global slow conduction. Reduced action potential duration
facilitates arrhythmia perpetuation but is not required. Up to 30% inhibition
of Is current has been introduced in various simulations.
Although most attempts resulted in unstained wavefront perpetuation, by
carefully timing the two ectopic beats, the model was able to initiate
atrial fibrillation and sustain it for more than 40 seconds (limited only
by the simulation time). Figure 2 shows an example
of atrial fibrillation initiated in a tissue with 10% inhibition of the
Is
current, a propagation velocity of c = 30 cm/s, and a stimulation
protocol involving two ectopic beats S2 and S3
located in the high right atrium with an S1-S2 interval
of 350 ms and an S2-S3 interval of 225 ms. After
an initiation period with multiple wave breaks and wave front interactions
due to slow recovery fronts [8], a fibrillatory
state appears in the model.

|
|
(a) 1650 ms
|

|
|
(b) 1700 ms
|

|
|
(c) 1750 ms
|

|

|
|
(d) 1800 ms
|
Figure 2: Snapshots of atrial fibrillation (from 1650 ms to 1800
ms after basic sinus rhythm initiation). AF was initiated by applying two
ectopic beats S2 and S3 with a coupling interval
of 350 ms and 225 ms respectively. The propagation velocity is c = 30 cm/s
and the action potential duration is 260 ms.
In this example fibrillation last for about seven seconds and then is
converted spontaneously to sinus rhythm. Compared to the periodic pattern
of atrial flutter, atrial fibrillation is observed as multiple reentering
wavelets traveling randomly and interacting with each others. Fibrillation
involves both anatomically anchored and free spiral tips, thus being a
complex mixture of anatomic and functional reentries, clearly visible in
the PV region of Figure 2. The average rate of
atrial fibrillation in our model is 420 bpm.
Similarly to what we can observe in clinical experiments, most of the
simulated atrial fibrillations convert spontaneously to atrial flutter
or sinus rhythm. An example of initiation of atrial fibrillation followed
by a spontaneous conversion to atypical flutter is presented in Video
2, which shows 10 seconds of real time propagation. Initiation involves
a 3 ectopic beats protocol following a sinus rhythm depolarization, very
similar to the one used for Figure 2, in a tissue
also having similar properties (c = 30 cm/s, Is
inhibition of 10%). After 2 seconds, a fibrillatory state is reached and
last for about 3 seconds. Finally, a spontaneous reorganization occurs
and stabilizes the remaining wavefront anchored to IVC and TV, thus leading
to a stable and periodic atypical flutter. The difference between the flutter
and fibrillation patterns is clearly visible in Video
2.
It is interesting to observe the number of wavelets during the initiation,
perpetuation and conversion of the atrial fibrillation described in Figure
2 (see Figure 3(a)). The stimulation protocol
induces one main wavelet having one of its tip anchored around SVC, while
its other free tip starts meandering. After a 1500 ms initiation phase,
multiple wavefronts are produced from the free spiral tip and spread in
the whole tissue, finally leading after 2500 ms to atrial fibrillation
with four to seven wavefronts. Episodes with few wavefronts sometimes happen
during fibrillation: two of such episodes can be seen in Figure
3(a) around 5000 ms, where only one single wavefront remains for few
milliseconds, but because of inhomogeneities in tissue repolarization due
to the previous fibrillatory state, multiple wave breaks rapidly lead back
to fibrillation instead of sinus rhythm. The simulations also confirm the
multiple wavelets hypothesis by Moe [9], which states
that fibrillation is maintained by the presence of at least four independent
wavelets.
Video 2: Initiation of atrial fibrillation and spontaneous conversion
to atypical flutter. The video is displayed real time and the total duration
of the simulation is 10 seconds. The propagation velocity is c = 30 cm/s
and the action potential duration is 260 ms.
(The VLC Media Player is recommended for viewing the video.)
In addition to the number of wavelets, the amount of excited tissue,
defined here as a transmembrane potential Vm > -70 mV,
has also been computed and is shown in Figure 3(b).
The initiation phase covering the first 2500 ms of simulation is characterized
by important variations in the percentage of excited tissue . This reflects
the global tissue behavior driven by a unique reentrant wavefront. The
periodicity of this wavefront anchored around SVC is responsible for the
periodic variation of the amount of excited tissue. Then a significant
drop of these variations can be observed during the first fibrillatory
episode characterized by a high number of wavelets (from 2500 ms to 3500
ms). This can be explained by the fact that the periodic behavior is replaced
by multiple simultaneous depolarization and repolarization processes each
related to an independent wavelet, globally leading to significantly reduced
variations in the amount of excited tissue (stabilized between 50% to 60%).
As the number of wavefront decreases, an augmentation of the variations
is observed. It can be noted that between 5500 ms and 6500 ms, there is
a gradual reorganization of the fibrillation process that will finally
lead to sinus rhythm.

|
|
(a)
|

|
|
(b)
|
Figure 3: Initiation of atrial fibrillation and spontaneous conversion
to sinus rhythm: (a) development of the number of independent wavelets,
(b) development of the percentage of excited tissue (Vm > -70
mV) . The stimulation protocol is described in Figure
2.
Conclusions
We have built a virtual atrium using validated
membrane channel physiology and conduction features as described in physiological
experiments. As the size and conduction velocity are within realistic values,
the compromise made so far in this model is that the atrial walls are considered
to be a single cell layer only. With few exceptions, this is acceptable
for the study of atrial arrhythmias, while it would not be the case on
the ventricular level. Another limitation of our model is that the whole
atrial tissue (except the anatomical holes) has uniform properties (action
potential duration and propagation velocity). Future work will consist
in the development of a more accurate structure and geometry and the introduction
of anisotropy and heterogeneity based on data from research in basic electrophysiology
and clinical experiments on atrial arrhythmias.
Simulations of up to 40 seconds of real time have been performed, while
in the literature, results in realistic models with a high number of spatial
nodes are limited to about one second [1]. Functional
and anatomic reentries were initiated only for appropriate basic conditions,
such as slowed conduction or reduced action potential duration. During
our virtual electrophysiological experiments, we have been able to initiate
atrial flutter-like reentry showing a single macro reentrant circuit with
a periodic pattern, as well as atrial fibrillation. The number of independent
wavelets is comparable to those observed in humans [7,10].
These simulations allow us to observe in great detail how atrial arrhythmias
originate from the prematurely stimulated tissue and how they are perpetuated
or spontaneously converted to atrial flutter or sinus rhythm. One important
observation is that in our model atrial arrhythmias are a combination of
functional and anatomic reentries and therefore the geometry plays an important
role. Indeed, the effect of anatomy of the heart on the mechanisms of arrhythmia
is clearly visible in our experiments. In the mechanism of fibrillation,
we have observed that veins and valves have two contradictory effects:
on one hand they act as anchors for the waves [11],
having therefore a stabilization effect and determining the reentrant pathways
as well as the rate of arrhythmia. On the other hand they also induce multiple
wave breaks mainly in the PV region, thus destabilizing propagation as
shown in the snapshot sequence of Figure 2.
Pulmonary veins were known to be arrhythmogenic [12],
but from our experiments they also seem to play a major role in fibrillation
perpetuation because of their wave breaking and anchoring capacities. Another
critical region for the initiation of atrial arrhythmias is the isthmus
between IVC and TV.The transition from fibrillation to a periodic and more
stable reentrant wave is facilitated due to the anchoring phenomenon. It
seems that major anatomical obstacles like IVC, SVC and both valves play
a more important role in this anchoring process than smaller holes like
PV.
In conclusion, our model of human atria, even if it is based on simplified
assumptions, can reproduce atrial arrhythmias showing a similar behavior
than reported in clinical experiments. The advantage of the computer model
is that it can show details difficult to study in nature and the experiments
are reproducible. It will be used as a tool to study the potential and
theoretical impact of the following therapeutic strategies: anti-tachy
pacing, defibrillation, ablation and even pharmacological interventions.
References
[1] Henriquez, C. S. and Papazoglou, A. A., "Using
computer models to understand the roles of tissue structure and membrane
dynamics in arrhythmogenesis", Proc. IEEE, vol. 84, no. 3, pp. 334-354,
1996.
[2] Gray, R. A. and Jalife, J., "Ventricular
fibrillation and atrial fibrillation are two different beasts", Chaos,
vol. 8, no. 1, pp. 65-78, 1998.
[3] Beeler, G. W. and Reuter, H., "Reconstruction
of the action potential of ventricular myocardial fibers", J. Physiol.,
vol. 268, pp. 177-210, 1977.
[4] Luo, C. and Rudy, Y., "A model of the ventricular
cardiac action potential", Circ. Res., vol. 68, no. 6, pp. 1501-1526,
1991.
[5] Wu, J. and Zipes, D. P., "Effects of spatial
segmentation in the continuous model of excitation propagation in the cardiac
muscle", J. of Cardiovascular Electrophysiology, vol. 10, no. 7, pp. 965-972,
1999.
[6] Virag, N., Vesin, J.-M. and Kappenberger,
L., "A computer model of cardiac electrical activity for the simulation
of arrhythmias", PACE, vol. 21 (Part II), no. 11, pp. 454-466, 1998.
[7] Allessie, M. A., Rensma, P. L., Brugada,
J., Smets, J. L. R. M., Penn, O. and Kirchhof, C. J. H., "Pathophysiology
of atrial fibrillation", in Cardiac Electrophysiology From Cell to
Bedside. Edited by Zipes, D. P., and Jalife, J., Saunders, pp. 548-559,
1990.
[8] Courtemanche, M., "Complex spiral wave
dynamics in spatially distributed ionic model of cardiac electrical activity",
Chaos, vol. 6, pp. 579-600, 1996.
[9] Moe, G. K., "On the multiple wavelet hypothesis
of atrial fibrillation", Arch. Int. Pharmacodyn. Ther., vol. 140, pp. 183-188,
1962.
[10] Shilling, R. J., Kadish, A. H., Peters,
N. S., Golberger, J. and Wyn Davies, D., "Endocardial mapping of atrial
fibrillation in the human right atrium using a non-contact catheter", European
Heart Journal, vol. 21, pp. 550-564, 2000.
[11] Xie, F., Qu, Z. and Garfinkel, A., "Dynamics
of reentry around a circular obstacle in cardiac tissue", Physical Review
E, vol. 58, no. 5, pp. 6355-6358, 1998.
[12] Haïssaguerre, M., Jaïs,
P., Shah, D., Takahashi, A., Hocini, M., Quiniou, G., Garrigue, S., Le
Mourroux, A., Le Métayer, P. and Clémenty, J., "Spontaneous
initiation of atrial fibrillation by ectopic beats originating in the pulmonary
veins", New Eng. J. of Medicine, vol. 339, no. 10, pp. 659-666, 1998.

 Home
Current Issue
Table of Contents
Home
Current Issue
Table of Contents


