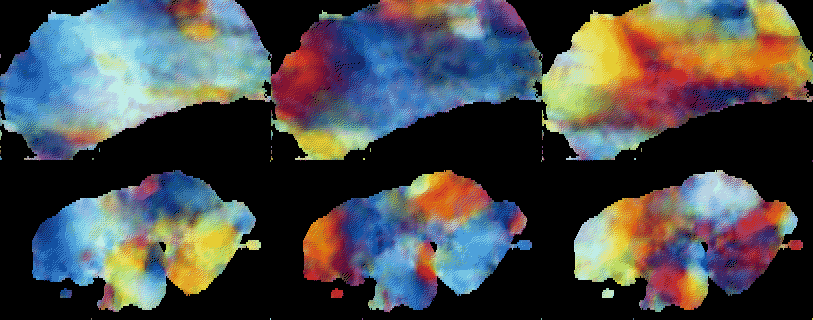|
Computational Biology of Propagation in
Excitable Media Models of Cardiac Tissue
A. V. Holden* and V. N. Biktashev**
* Computational Biology
Laboratory, School of Biomedical Sciences, University of Leeds,
Leeds LS2 9JT, UK
** Division of Applied Mathematics, Department of Mathematical Sciences,
University of Liverpool,
Liverpool, L69 3BX, UK
Correspondence: arun@cbiol.leeds.ac.uk
Abstract. Biophysically detailed
models of the electrical activity of single cardiac cells are modular,
stiff, high order, differential systems that are continually being updated
by incorporating new formulations for ionic fluxes, binding and sequestration.
They are validated by their representation of the ionic flux and concentration
data they summarise, and by their ability to reproduce cell action potentials,
their stability to perturbations, structural stability and robustness.
They can be used to construct discrete or continuous, one-, two- or three-dimensional
virtual cardiac tissues, with heterogeneities, anisotropy and realistic
cardiac geometry. These virtual cardiac tissues are being applied to understand
the propagation of excitation in the heart, provide insights into the generation
and nature of arrhythmias, aid the interpretation of electrical signs of
arrhythmia, to develop defibrillation and antiarrhythmic strategies, and
to prescreen potential antiarrhythmic agents.
Keywords:Cardiac action potential,
propagation, arrhythmia, re-entry, defibrillation
Introduction
The computational modelling of electrical activity in the heart has
provided a quantitative, detailed description of normal activity, and
is being applied to understand cardiac arrythmias, and to evaluate
methods for their control or prevention. Cell excitation is
described by stiff, high order systems of ODES that have and are being
obtained from voltage clamp experiments on single cells and membrane
patches. Cell models can be coupled to form tissue models, either in
discrete space (coupled ODE lattice models) or as partial differential
systems of the reaction diffusion type. These are heterogeneous, both
in the sense that different cell types can be intermixed in the same
tissue (\eg\ fibroblasts and pacemaker cells in the sinoatrial node),
and regional differences in cell parameters (say endo- to epicardial
changes in action potential shape prodiced by quantitative changes in
membrane ionic conductance parameters). Cardiac tissue is anisotropic,
with propagation faster along the fibre axis: homogeneous aniostropy
can be removed by coordinate transformation, but rotational anisotropy
cannot. Quantitative models for cardiac geometry and anisotropy exist
(for the canine ventricles), and are being developed (for the pig
and human atria). Thus the investigation of propagation and its
disorders is highly computational, and has developed in close parallel
with the avilablity of adequate computing power. Since cell models are continually being updated by the
incorporation of new results these computational models need to be
highly modular, so individual components can be unplugged and updated.
Modern cellular electrophysiology has provided quantitative descriptions, for
different types of cardiac muscle cells of different species, of membrane
ionic currents, pumps and exchange mechanisms that have been combined with
intracellular and extracellular ionic accumulation, depletion and sequestration
processes to form biophysically detailed models of membrane excitation
[1]. These models, in the form of high order (a large
number of state variables), stiff (time scales ranging from fractions of
a ms to hundreds of ms) differential systems may be integrated to produce
numerical solutions that reproduce currents seen under voltage clamp, or
membrane potential time series recorded from single cells. Models for cells
from different parts of the heart - the sinoatrial node, atrium, atrio-ventricular
node, Purkinje fibres and ventricular cells - have different action potential
characteristics, generated by quantitatively different but qualitatively
similar mechanisms. Any single cardiac muscle cell can be modelled by a
system of ion-selective conductances with voltage-dependent activation
and inactivation processes, ionic pumps and exchangers, together with intracellular
and extracellular ionic sequestration, depletion and binding as differential
system:
dV/dt = -I(V,gn )/C
dgn /dt = G(V,gm)
n,m = 1,...,N
where C is the cell capacitance, V is the transmembrane voltage, I the
transmembrane current, the variables gn that describe the state
of a cell (gating variables for the different ionic channels, ionic concentrations
in different compartments) and the functions G describe their dynamics.
The apparent simplicity of this description hides its complexity: N can
be large (e.g. N=17 for the guinea pig ventricular cell models [2]
used below, and it also contains a large number of parameters (e.g.
maximal conductances, ionic concentrations, reversal potentials), some
of which are based on experimental estimates, and some of which have been
chosen to satisfy some constraint. For the cell model to have a stable
resting potential (a solution such that dV/dt =0) the resting state is
electrically neutral i.e. charge entry and exit via channels,
pumps and exchangers is balanced. However, an electrically model need not
be chemically neutral - unless the entry and exit rates of each ionic species
are balanced there will be slow changes in intra- and extra-cellular ionic
concentrations with time. Cell models and their parameter values have been
constructed primarily from electrophysiological experimental data from
different sources, obtained by different methods, and usually obtained
by protocols with a time scale of one to a few hundred ms. They are usually
not chemically neutral and so the valid time scale for cell models (and
tissue models derived from them) is only of the order of seconds: over
longer time scales there are slow changes in the variables representing
concentrations that produce artefactual behaviours.
The variety of different cell models, and the alternative models for
the same cell type, combined with their common basic structure, suggests
a modular approach in which a particular cell model is specified by a set
of modules that represent the ionic transfer mechanisms (ion-selective,
voltage dependent channels; pumps and exchangers), together with binding
and sequestration mechanisms (e.g. Ca++- binding by phospholamban,
calmodulin). Each of these mechanisms corresponds to a protein or protein
complex, and so will be able to be mapped onto the proteome. Each mechanism
has its associated magnitude (corresponding to its membrane density or
intracellular concentration), and dynamics, represented by a normal range
of parameter values. These parameter values differ between different models,
and can be modified to represent the effects of changes in the cellular
environment (e.g. temperature, via the Q10 of
rate coefficients), pharmacological agents (e.g. see [3] for examples of channel blockers) or mutations in genes expressed
as cardiac channels (e.g. see
Meander in LQT syndromes below).
Thus a specific model of a normal or abnormal cell can be assembled from
a set of modules and parameters, as in the Oxsoft [4]
package. As yet, there is no public domain package that would provide cellular
cardiology with an equivalent to what GENESIS [5 ] provides
cellular neurophysiology.
The excitation equations for all cardiac muscle cells are high order
and complex, with a large number of variables and parameters. Vertebrate
axonal excitation equations have less than four dynamic variables controlling
two conductances, while cardiac excitation equations typically have about
20 dynamic variables controlling about a dozen conductances. From the computational
viewpoint, this raises practical problems, as there are few published models
that are without typographical errors, and so ensuring that a program actually
codes a given model and accurately specifying that model, is not as straightforward
as it should be. From the functional viewpoint, the complexity of cardiac
excitation may just be an illustration of the "baroque" nature of biology,
as for excitability and autorhythmicity only two variables are necessary,
and for the rate dependent changes in action potential duration that are
mapped as electrical restitution curves only three variables are required.
However, the mechanisms of cardiac excitation has not been sculptured but
have evolved, and the complexity may give cardiac excitation a robustness
to changes in parameters. In spite of homeostatic mechanisms, life threatening
changes in the internal environment , such as fever, changes in pH, and
osmolarity do occur as part of the trials of life, and the complexity of
the cardiac excitation mechanisms might provide a robustness of behaviour
- a persistence of sinus rhythm- in the face of these large fluctuations
in parameters.
A virtual tissue can be constructed by coupling together cell models,
either in a discrete representation, as a lattice of coupled cells, or
in a continuous representation, as a system of partial differential equations
of the reaction-diffusion form, where the "reaction" term represents the
nonlinearities of membrane excitation and the diffusive term the electrotonic
spread of potential with distance through the cardiac tissue.
Such a virtual tissue can be used to understand the physiology of propagation
in cardiac tissue - for example, propagation during the normal sinus rhythm
is often from tissue with longer to tissue with shorter action potential
duration, as from the centre to the periphery in the sino-atrial node
[6] and
from the endocardial to epicardial surfaces of the ventricular wall
[7],
so the depolarisation wavefront propagates "orthodromically", while
the repolarisation waveback collapses "antidromically". A consequence is
that re-entrant propagation is prevented.
Failure of the rhythmic pumping of the heart produced by the arrhythmias
of ventricular tachycardia and fibrillation is not only a major cause of
death, but is a terminal event in almost all non-violent deaths. Most of
these deaths are premature, both in the sense that the probability of occurrence
of a vascular insult to the myocardium triggering a lethal arrhythmia can
be reduced by appropriate dietary and activity regimes, and that potentially
lethal arrhythmias can be terminated by defibrillatory interventions if
they are applied soon enough. Virtual tissues can be used to understand
the mechanisms of initiation and persistence of arrhythmias, to explore
the phenomenology and methods of defibrillation, and to design or prescreen
antiarrhythmic agents.
Below we illustrate the use of virtual cardiac tissue in understanding
and controlling re-entrant ventricular arrhythmias, by considering case
studies of ventricular re-entry, LQT syndromes, resonant drift as a strategy
for low-voltage defibrillation, the behaviour of weakly excitable tissue,
bidomain models and virtual electrode effects in defibrillation, and three
dimensional aspects of ventricular fibrillation.
Methods
Cardiac tissue is spatially extended, and the description of propagation
and its disorders requires models of cardiac tissue as an excitable medium.
The study of wave propagation in `reaction-diffusion' models of excitable
media, i.e. considering tissue as a continuous syncytium, has already
contributed to the understanding of many phenomena related to cardiac electrophysiology
and has been done mostly in one-, two- and three dimensional models of
excitable media with simplified kinetics. Extensive exploration of two-dimensional
media or three-dimensional media with biophysically realistic kinetics
has only recently become possible. The study of three-dimensional cardiac
tissue models with realistic kinetics and anisotropy is only just becoming
possible. The problem with biophysically realistic models is their stiffness,
i.e. wide range of characteristic time and space scales: from tens
of microseconds to hundreds of milleseconds and from tens of micrometers
to centimetres, thus the computational cost of straightforward approaches
is enormous. We are developing multigrid or restructurable grid schemes
to reduce this load.
Granularity
`Reaction-diffusion' approaches to cardiac tissue cannot in principle
describe some experimental phenomena. One example is the anisotropic vulnerability
[8] , the phenomenon of different minimal period of
propagating waves depending on the direction of propagation, which is impossible
in continuous homogeneously anisotropic reaction-diffusion system (it is,
however, explainable in the bidomain theory, see below). An obvious way
to allow for this sort of phenomena is to consider each particular cell,
i.e. describe the tissue in terms of coupled ordinary differential
equations (CODE) as in [9] rather than partial differential
equations (PDE). In certain situations this can be avoided by using phenomenological
interval-velocity relationships accounting for the cellular structure [10];
this approach deserves further study.
Anatomy
Digitized anatomical data describing the canine heart (ventricles) including
fibre orientation are available [11], and such models
have already been used in pilot simulations with simplified kinetics. Early
studies of propagation in realistic tissue geometries in mesoscopic [12]
and macroscopic [13 14]
scales were all done with simplified reaction-diffusion models. Incorporating
``rotational anisotropy'' within this approach does not meet any serious
difficulties, as all that is required is using a conductivity tensor instead
of isotropic diffusion of potential. To date computations of propagation
in anatomically realistic models of cardiac tissue have been in a static
geometry; and propagation phenomena in a moving medium is beginning to
be approached, using phenomenological models [15 16
].
Bidomain equations of cardiac tissue
Cardiac tissue can be considered as consisting of two domains: the interiors
of the cells, which are electrically connected by Ohmic gap junctions,
and the common exterior, the two domains being separated by the cell membranes,
where the nonlinear nature of cardiac excitability is localised. The currently
prevailing viewpoint is that the distribution of the electric potential
in each of the domains is normally more or less smooth. This enables averaging
of the conductivity properties within each of the domains over the cellular
scale. This averaging leads to the description of the excitation propagation
in terms of a PDE system, which can be written as a system of local equations
for the transmembrane voltage E and local excitation variables (channel
gates, ionic concentrations etc), and elliptic equation for the extracellular
potential 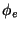 : :
:
is transmembrane current, gi are local variables
and  is cardiocyte surface/volume ratio.
is cardiocyte surface/volume ratio.
The key parameters of these equations are the conductivity tensors σi
and σe
,
of the two domains, interior and exterior. If the corresponding components
of the two conductivity tensors relate to each other by a constant factor,
the elliptic equation degenerates, and the system is reduced to the parabolic
equation, the "monodomain" or "cable" theory (which can be also obtained
in the limit if one of the conductivity tensor is infinitely large) -
see [17 18 19 20]
and references therein. In general, both the equations for external
potential and transmembrane voltage (or equivalently, external and internal
potentials) should be solved simultaneously, and this difference from the
monodomain theory provides specific features of excitation propagation,
like the non-elliptic shape of the waves from a point source and anisotropic
dispersion (velocity-rate) relationship. Another, qualitatively important
feature of the bidomain equations is that they describe the relationship
between external electric field and the distribution of the transmembrane
potential, which is important both in the interpretation of electrocardiographs
and for electrical stimulation and defibrillation technology. Computational
approaches for the bidomain equations vary, and include e.g. spectral
methods [18], method of Green functions [19]
and alternating directions [20]. The first
two are applicable in the case of spatial uniformity of the conductivity
tensors (in particular, a constant direction of the fibres) and are therefore
of limited interest. The AD method is applicable to the regular rectangular
grids and can therefore only be considered as a starting point, or as an
interim procedure in a multigrid approach. An appropriate iteration procedure
for resolving the elliptic equation, that can be reformulated as a sequence
of explicit steps, and can therefore be applied to the multigrid tree and will
allow parallelization (see below). Due to the disproportion of characteristic
times, the description of the distribution of transmembrane voltage and
fastest membrane variables over the membrane of one cell can be successfully
reduced to a low-dimensional O.D.E. [21] , and so
the effects of membrane parameters on defibrillation thresholds can readiliy
be computed.
The trimmed-tree multigrid approach
In computation with regular grids, the time and space resolution are
determined by the temporal and spatial scales of the excitation front,
of the order of 1 msec and 1 mm respectively, which require corresponding
computational steps to be at least of order of magnitude less, whatever
the requirements of precision or stability. On the other hand, this resolution
is required only at the fronts, and away from the fronts both time
and space variations are quite smooth and much larger steps could be acceptable.
The idea is to use small steps at the fronts and large steps away from
it. Since the geometry and even the topology of the fronts is varying not
only from experiment to experiment, but even in the course of one experiment
(and, in a sense, such topological deformations are one of the most principal
issues of the theory, as they correspond to birth or death of re-entry
waves), all the computational approaches which make any assumptions on
these fronts are not acceptable. We use the idea of a trimmed quadtree
representation, widely used in image representation technology. The 2D
medium is split onto square cells, and if the variation of the dynamic
field within one cell exceeds a certain value, this cell is split onto
four children cells. This process is repeated iteratively until variation
of the field within every cell is less than the adopted criterium. As the
front propagates, the cells it approaches should split and the cells behind
it may join together again. This idea obviously extends to 3D.
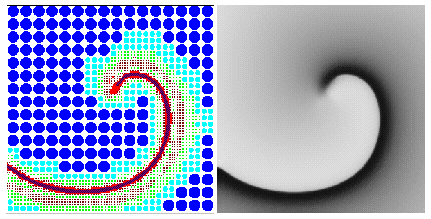
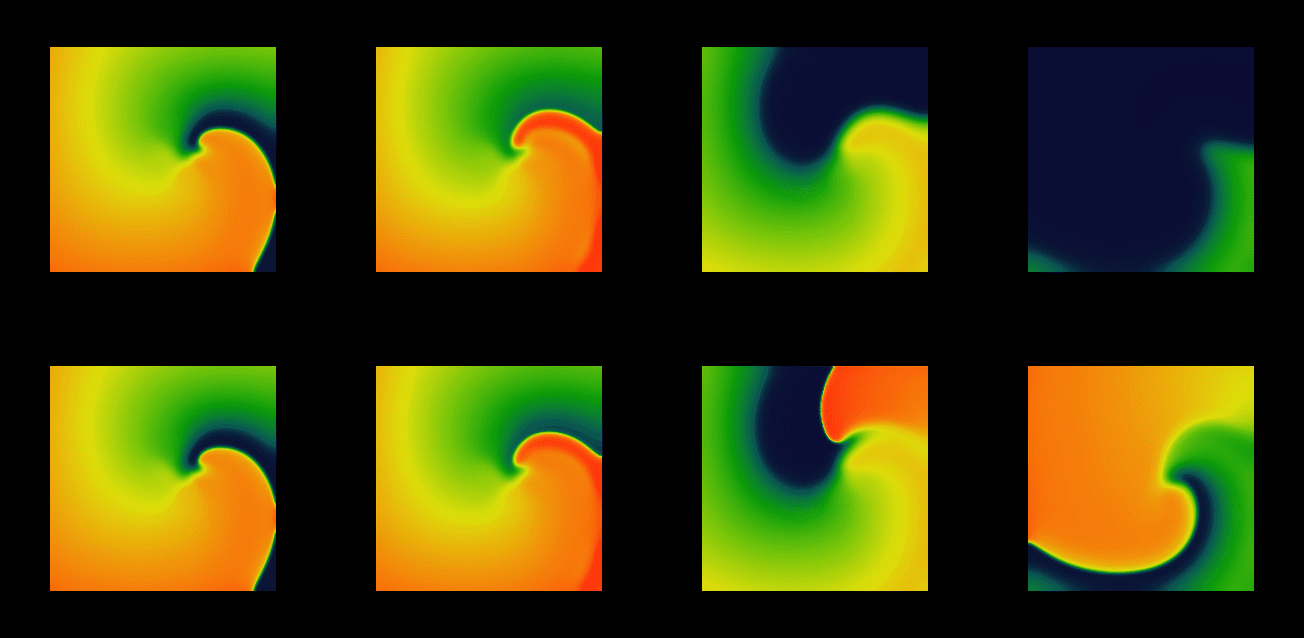
Figure 1: A multigrid with 6 resolution layers (left) built to
approximate a stiff spiral wave wave solution (right). The number of nodes
is 30 times less than that required by a regular grid with the same accuracy.
This approach is illustrated in Figure 1. In practice, we use not a
tree, but a forest, with roots arranged in a regular grid of small size
(16x16 for the example shown on the figure), implemented as a dynamic structure.
For models with biophysically realistic kinetics,
the main computational load (typically no less than 95%) is on the local
step, which can be performed absolutely independently for all computational
cells, thus making this approach perfectly parallelizable, with the maximal
potential degree of parallelization being the number of computational cells.
As for the non-local step, it is also parallelizable if explicit schemes
are used. The stiffness of cardiac excitable kinetics imposes severe restrictions
on the time step at the front anyway, so explicit schemes for the non-local
part may be acceptable in many practical situations, including bidomain
equations, if relaxation method can be accepted for resolving the elliptic
equations.
Results
Propagation phenomena in one-dimensional virtual tissues
The reaction-diffusion equation in one dimension has a spatially uniform
solution, corresponding to resting tissue, and can support solitary wave
and wave train solutions. The solitary travelling wave solution propagates
at a velocity proportional to the square root of the diffusion coefficient,
and so the diffusion coefficient can be chosen to give appropriate length
and velocity scaling, or can be obtained from estimates of cell to cell
coupling conductance. The velocity of travelling wave solutions is rate-dependent.
Two travelling wave solutions meeting head on collide and annihilate
each other; this destructive interference results from the refractory period
of the travelling waves. Supra-threshold stimulation at a point in a uniformly
resting one-dimensional model produces a pair of travelling wave solutions
that propagate away from the initiation site. The initiation of a single
solitary wave in a one-dimensional ring provides a computationally simple
model for re-entry; such unidirectional propagation can only be produced
in a homogeneous one-dimensional medium if the symmetry is broken, say
by a preceding action potential. The vulnerable window is the period after
a preceding action potential during which a unidirectional wave in a one
dimensional medium can be initiated; stimulation during the vulnerable
period in the wake of a plane wave in a two-dimensional medium would initiate
a pair of spiral waves.
The length of the vulnerable window increases with stimulus intensity
and with the length of the stimulated tissue (the electrode size). If the
effects of pharmacological agents or pathological processes (ischaemia,
acidosis) can be expressed as changes in the excitation system then estimating
the vulnerable window provides a means of quantifying the pro- or anti-arrhythmogenic
effects of these changes. An increase in the size of the vulnerable window
increases the likelihood of re-entry being triggered: this is found for
Na+-channel blockers [22 ] and so can account
for the pro-arrhythmogenic effects of the agents used in the CAST trials
[23].
Re-entry in a two-dimensional ventricular virtual tissue
The generalisation of a solitary wave and a wave train in a two dimensional
medium is a plane wave and plane wave train. Since cardiac cells are cylindrical,
and organised in sheets, propagation in cardiac tissue is anisotropic,
with the velocity being faster parallel to the fibre axis. In homogeneous
anisotropic cardiac tissue the "wavelength" of the action potential changes
with its direction of propagation. For isotropic virtual tissue repetitive
focal excitation will generate a circular wave train, the ellipsoid propagation
pattern seen in real cardiac tissue can be produced simply by a simple
co-ordinate transformation. The normal velocity V of a wavefront is also
dependent on its curvature k: V = V0 -Dk, where
D is the effective diffusion coefficient. The dependence of velocity on
rate, and on curvature, allows rotating spiral wave solutions.
A spiral wave in a two-dimensional, homogeneous, isotropic medium provides
a model for re-entry. Spiral waves rotate around a central area of conduction
block, or core, and may be characterized by their period of rotation, size
of core, and movement of the tip of the spiral. At any specified instant
in time the spiral wave has a location (given by the position of its tip),
and a spatial orientation of rotation phase. The tip can rotate rigidly
around a circular core, whose radius increases as excitability decreases,
or meander bi-periodically [24 25 26].
For isotropic atrial virtual tissue, the spiral wave initially rotates
rigidly, around a circular core, with a period of 73 ms [27].
As the spiral wave ages the period increases to 84ms over 5 s, and
the size of the core increases and the tip begins to meander [28].
The period of the spiral wave is close to the period of atrial flutter.
For isotropic ventricular virtual tissue, the spiral wave illustrated
in Figure 2 rotates with a period of initially 170 ms, that decreases over
a second to 100-110 ms. The motion of the tip is not circular, but meanders,
moving by a jump-like alternation between fast and very slow phases, with
about five jumps per full rotation of the spiral.
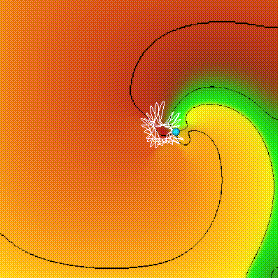
Figure 2: Spiral wave solution for ventricular tissue model.
The intersection of two isolines (for V = -10 mV, and the Ca++-inactivation
variable f = 0.5) defines the position of the tip (blue ball) whose trajectory
appears as the white curve. [2]
The multi-lobed meander of the ventricular tissue model we use, that
has extended linear segments separated by sharp corners, has been accounted
for in terms of the two time courses of the two principal depolarising
currents. Propagation of the re-entrant spiral, or of a wave around an
extended linear obstacle, alternates between being driven predominantly
by sodium and calcium currents. Modification of the ratio of the time courses
of these two currents can extend the near- linear segments of meander.
If these near-linear segments are of the same length as the distance to
an inexcitable boundary of the medium then the meander of a re-entrant
wave would lead to its self termination by moving its tip to an inexcitable
boundary. This mechanism might provide an explanation for ventricular tachycardia
that manifests itself as syncope, and for episodes of self-terminating
fibrillation observed when the electrocardiogram is being continually monitored,
as in intensive or coronary care units [29].
LQT syndromes: meander, self-termination and lethality.
Inherited LQT syndromes are associated with increased risk of re-entrant
arrhythmia and result from mutations in genes expressed as cardiac Na+-
and K+ - channel subunits . These mutations prolong ventricular
action potentials and produce long Q-T (LQT) intervals in the electrocardiogram.
Arrhythmias occur more frequently in patients with LQT1 and LQT2, associated
with mutant K+- channels, yet are 5 times more likely to kill
patients with LQT3, associated with mutant Na+- channels [30].
We interpret this finding as a greater likelihood of self-terminating
re-entry in LQT1 and LQT2. The relative meander of re-entrant sources in
these three phenotypes is consistent with clinical outcome, and illustrates
that computational functional genomics can provide insights into the whole
organ consequences of genetic abnormalities. The specific gene mutations
each associated with an LQTS have been identified [31 32].
Of these, LQT1 is a mutation of the KVLQT1 and/or hminK genes which
reduces the magnitude of the slowly activating delayed potassium current
IKs , LQT2 a mutation in the HERG gene which reduces the magnitude
of the rapidly activating potassium current IKr, and LQT3 a
mutation in the SCN5A gene which prevents complete inactivation of the
sodium current INa. Episodes of arrhythmia, identified by syncope,
documented tachyarrhythmia, or sudden cardiac death, occur most often in
patients with LQT1 and least often in patients with LQT3. However, the
incidence of lethal arrhythmias is five times greater in patients with
LQT3 than in patients with LQT1 or LQT2 . Episodes of arrhythmia in patients
with LQT1 and LQT2 are therefore more likely to self-terminate than those
in patients with LQT3. Many LQTS arrhythmias show the characteristic waxing
and waning in the electrocardiogram (ECG) that is classified as torsade
de pointes by cardiologists. We assume that LQTS arrhythmias, once
initiated, are sustained by a propagating re-entrant wave rotating around
a moving core, and that this single wave then can break down into the multiple
waves of fibrillation. Meander occurs in homogeneous isotropic and anisotropic
media, in heterogeneous media the meander is accompanied by drift. Re-entrant
waves can be extinguished when their core either drifts or meanders to
an inexcitable boundary. Both meander and self-termination of re-entrant
waves have been observed experimentally. In cardiac tissue a meandering
and/or drifting re-entrant wave can be pinned by a discrete anatomical
obstacle, such as a blood vessel. Self-termination of an unpinned re-entrant
wave is more likely if the extent of meander is greater, because the core
is more likely to move to a boundary between heart muscle and inexcitable
connective tissue. Our model of normal myocardial tissue had an action
potential duration (APD) measured at 90% repolarisation of 153 ms for plane
waves when paced at a cycle length of 1000 ms. In the models of LQTS myocardium
APD was prolonged by between 14 and 19%. The cellular restitution curves
for simulated LQT2 and 3 are monotonic and similar to that for the normal,
shifted upwards towards longer durations, while the restitution curve for
LQT1 shows evidence of supernormal action potentials at very short intervals.
The meander of the core in simulated LQT1 (Figure 3 right) is both greater
in extent and more irregular than in LQT2 and LQT3 (Figure 3 left) In particular,
it has extended linear components, that in anisotropic tissue would be
up to three times longer. It is these fast, linear components of meander
that increase the likelihood of the core reaching an inexcitable boundary.
In LQT2, the tip trajectory is similar to normal myocardium although the
corners are smoother. In LQT3 , the trajectory is again similar to normal
myocardium, except that the corners are sharper. Apart from the very small
differences in action potential duration, the major difference between
the LQT1, LQT2 and LQT3 simulations was the biphasic restitution curve
for LQT1. We conclude that in LQT1 the biphasic restitution curve exaggerates
the alternate fast, (INa driven) and slow (ICa driven)
meander cycles, leading to an increased meander in an isotropic medium
that would be amplified in a heterogeneous and anisotropic 3-D ventricle
to increase the likelihood of self terminating arrhythmias. The 5-fold
increased meander seen illustrated in Figure 3 is consistent with the increased
likelihood of self-termination for LQT1 as compared to LQT3 tachyarrhythmias
[33].
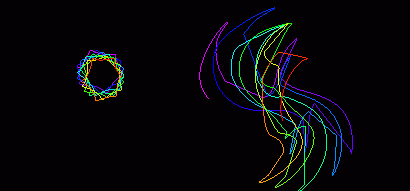
Figure 3: Computed spiral wave tip trajectory 1-2 s after initiation
by the phase distribution method in homogenous isotropic LQT 3 (left) and
LQT1 (right) virtual tissues, medium size 30 by 30 mm [33]
Resonant drift as a potential low-voltage method of defibrillation
A major cause of sudden death is the formation of a re-entrant wave
of excitation in the ventricles of the heart, that prevents the rhythmic
beating of the heart and its ejection of blood. In such a re- entrant wave
excitation propagates through the heart muscle, repeatedly re-invading
the same tissue; this re-entry can break down into ventricular fibrillation.
A spiral wave can be forced to move by a spatially uniform, time periodic
perturbation of appropriate frequency. Small amplitude, spatially uniform
repetitive stimulation can be used to produce directed movement of a rigidly
rotating spiral wave, if the period of stimulation is equal to the period
of the spiral wave rotation (resonant drift). If the stimulation period
is close but not equal to the rotation period of the spiral a circular
drift is obtained. If the stimulation period is fixed, this drift is strongly
influenced by medium inhomogeneities [34]. Resonant drift
in the location of a spiral occurs when the frequency of perturbation is
the same as the frequency of rotation of the spiral.
In principle, resonant drift under feedback control could provide a
means of eliminating re-entrant activity in cardiac tissue [35].
This contrasts with current methods of defibrillation, which use single,
large amplitude, shocks, that, although usually effective, does cause damage
to the heart muscle. The potential application is the market for "intelligent"
implanted cardiac defibrillators, trans-oesophageal atrial defibrillation,
and open chest defibrillation after fibrillation has been induced to allow
cardiac surgery. This will only be practical only if any re-entry is eliminated
within a reasonable time, say less than 30 s, and estimation of the velocities
of the directed drift that can be achieved by resonant drift is important
in assessing its feasibility as a means of controlling re-entrant arrhythmias.
Such a drift has been observed in reaction-diffusion model of rabbit
atrium based on Earm-Hilgemann-Noble kinetics [27],
as it initially generates rigidly rotating spiral waves. An appropriately
timed perturbation of 15% of the amplitude of the single shock defibrillation
threshold produces a directed motion with a velocity of about 0.4 cm/s,
and so resonant drift under feedback control could be used to eliminate
a spiral wave from the atrium within approximately 10 s, and so is a feasible
approach. This is an alternative to the proposed use of chaos control techniques
[36].
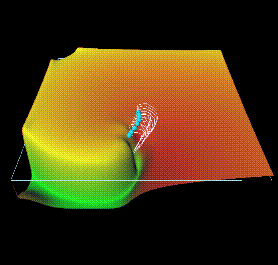
Figure 4: Perturbations applied at the same phase of each rotation
produce a directed drift of the tip of the spiral wave solution of Figure
2; the tip trajectory is the white line; the voltage
is represented by a vertical displacement.

Figure 5: Tip trajectory for ventricular virtual tissue under
feedback controlled, resonant driving. When the wavefront of the spiral
wave reaches a recording site at the bottom left hand corner, a 2 ms, 4V/s
depolarising perturbation was added after a fixed delay. Each trajectory
is for a different delay, corresponding to a different phase of the spiral.
All trajectories start in the centre, move towards the boundaries and annihilate.
The dots mark points on the trajectories corresponding to the moments of
stimulation.
In the ventricular virtual tissue, even in the absence of inhomogeneities,
the instantaneous frequency of the spiral is always changing, because of
the meander and so a pure resonant drift is not observed at any constant
frequency. The resultant motion is a nonlinear interaction between the
pattern of meander and the motion produced by the perturbations. The directed
motion of resonant drift is much more robust if instead of choosing a fixed
frequency, some kind of feedback is used to synchronise the stimulation
with the spiral wave rotation [35]. Such feedback control
can provide the stable resonant drift in the ventricular virtual tissue
model [2]. Figure 5 shows four tip trajectories produced
by repetitive stimulation applied at four different fixed delays after
the wavefront reached the bottom left corner. The delay determines the
initial direction of drift. A repetitive perturbation of 15% the amplitude
of the single shock defibrillation threshold produces a directed motion
with a velocity of about 0.4 cm/s.
Using simpler models, either the FitzHugh-Nagumo partial differential
equation, or its simplification for rapid computation for long times or
in 3-dimensional space, or a kinematic description of wave front motion,
or ordinary differential equation normal forms for the dynamics of meandering
spiral waves, the effects of boundaries, obstacles, and meander on the
near-resonant/resonant induced motion of spiral waves can be explored.
These studies provide a broad framework, within which some of the behaviours
seen are relevant to the control of re-entrant waves in cardiac tissue.
[37] uses simple FitzHugh-Nagumo equations to explore
the effects of inexcitable boundaries, electrode position, and inexcitable
obstacles, on resonantly induced motion of spiral waves in circular and
annular media, where the radius of the medium is of the same order of magnitude
as the spiral wavelength. An objection to the application of resonant drift
to heart muscle is that local inhomogeneities, like blood vessels, can
trap moving spirals; different feedback methods are used to overcome this
trapping.
Weakly excitable media
We have developed the kinematic approach to spiral motion , as distinct
from the eikonal approach (ie considers movement of the broken end of the
front of the wave, as opposed to the tip, defined by where wave-front and
-back meet [38]), and considered behaviours in weakly
excitable media close to Winfree's [39] rotor boundary,
where spiral waves fail to propagate even though plane waves can. This
leads to a more general kinematic approach, within which the approaches
of Davydov, Zykov and Mikhailov form a special case. The relevance of this is
that it takes the kinematic theory close to the region of reduced excitability/shortened
action potential duration that is characteristic of ischaemic tissue, and
so provides the beginnings of a theoretical framework within which numerical
simulations of re-entrant wave initiation and stability in models of ischaemic
tissue can be understood. We have applied this theory to the drift of the
spiral waves due to inhomogeneity of medium properties. Both the inhomogeneity
of the medium properties and the drift of the spiral/scroll waves are considered
as important factors of the fibrillation.
Qualitative features of meander
Mathematical aspects have been taken further, by using a general theoretical-group
approach to explain the main qulaitative features of meander of spiral
waves in the plane, based on the space reduction method to separate the
motions in the system into the superposition of those along orbits of the
Euclidean symmetry group, and those across. The system of ODEs governing
tip motion was obtained, and a derivation of the Barkley normal form/model
system for bifurcation from rigid to biperiodic rotation presented [24].
This approach was extended in [40] to account for hypermeander
as a chaotic attractor in the quotient system with respect to the Euclidean
group. Such an attractor should lead to motion of the tip of the spiral
analogous to the motion of a Brownian particle, with the mean square displacement
of the tip growing linearly at large times, and so leading to self-termination
of re-entry in a restricted medium.
Extracellular fields, bidomain models and the virtual electrode effect
A biophysical problem with the stiff, high-order, reaction-diffusion
models of cardiac tissue is that the effects of external voltage gradients
are not considered appropriately, as the tissue is treated as a continuum,
not cells embedded in extracellular fluid, and if an external field is
to influence a cell it must have different effects where it enters and
leaves the cell. For anisotropic tissues a bidomain approach is sufficient
for investigating propagation, so in place of one PDE there are two coupled
PDEs. However, for treating the effects of externally applied voltage fields,
each cell needs to be treated as a spatially extended object. A simplification
of this problem is given in [21], and so defibrillation
thresholds, and the effects of pharmacological agents on them, can be computed.
The above results about resonant drift were for external perturbations
modelled as an additional current in the equation for the transmembrane
potential, with an explicit time dependence. This is easy for numerical
simulation, but does not correspond to real situation, where the defibrillating
voltage current is not applied across the membrane, but imposed extracellularly.
Therefore, the above results are not directly comparable to experimental
data. Specifically, this concerns the values of amplitudes of the stimuli,
measured in mV/ms or nA/cell , which have little and indirect relation
to experimental values of V/cm or mA/cm2. This is not a matter
of mere rescaling, by estimating how much of the external current actually
penetrates the cell membrane, but is a more fundamental difference in biophysical
mechanisms of action of this current onto the cell, since the same current
will cross the membrane of the same cell at the same time in different
directions in different parts of the membrane, and thus will always have
both depolarising and hyperpolarising actions on the cell as a whole. So,
the amplitudes of the above numerical results may be interpreted, at most,
only qualitatively and in units relative to something that is also experimentally
measurable, e.g. defibrillation threshold (DFT).
An absolute quantitative estimation of DFT can be obtained by a quantitative
theory of the interaction of extracellular current with membrane excitation
processes. This has been applied to the ventricular virtual tissue, and
has led to the estimation which is, at least in the order of magnitude,
comparable to experimental values.
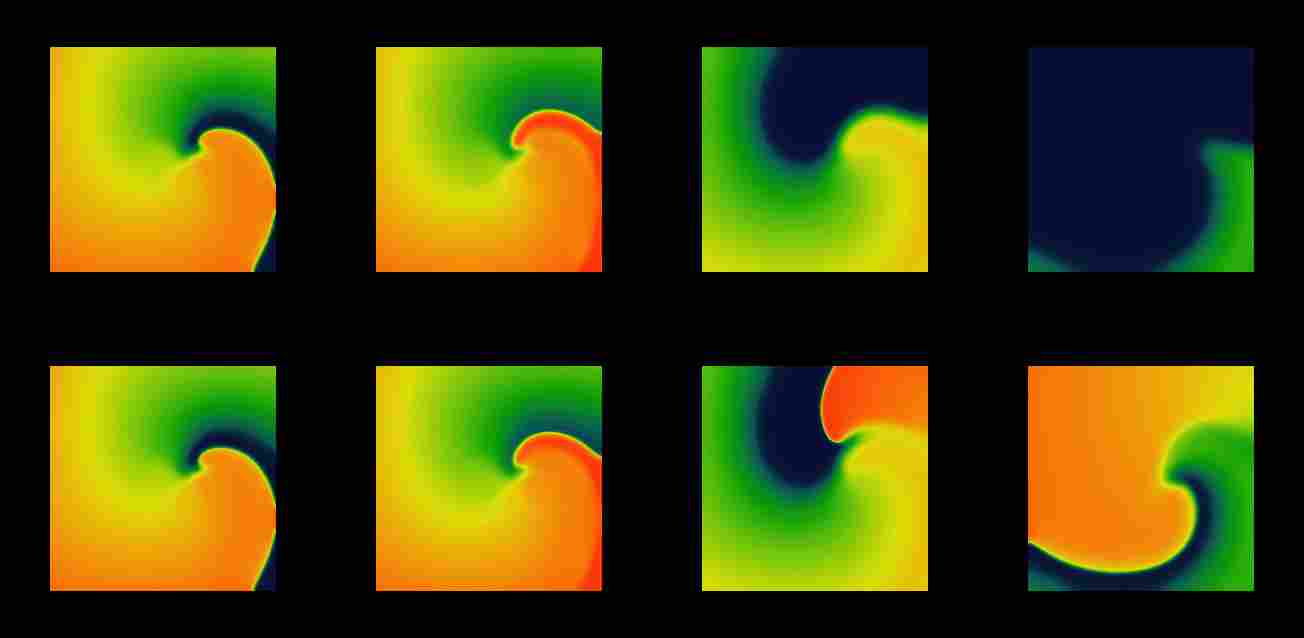
Figure 6: Snapshots from movies of suprathreshold (above, and
subthreshold (below) defibrillation by a spatially uniform depolarising
current pulse of a spiral wave as in Figure 1. Time moments are chosen
0, 3, 40 and 100ms (left to right) measured since the beginning of the
stimulus.
The stimulus has both depolarising and repolarising effects, and in
the region ahead of the front the depolarisation effect overbalances the
hyperpolarisation, and the front jumps forwards. The later evolution depends
on how far the wavefront jumped. If the stimulus was above the threshold
for defibrillation ( upper row of Figure 6), the front advances to the
region where the tissue has not recovered yet, and the antegrade propagation
is not possible. Hence, the front retracts, i.e. begins to collapse
backwards, and the excited region shrinks until it vanishes, as the depolarising
wavefront moves backwards and the repolarisation waveback carries on moving
forwards.
A smaller (subthreshold) shock will produce a smaller advance in the
position of the front and thus allow the possibility for it to recover
its forward propagation. This possibility depends on two factors, the refractory
state of the medium and the front curvature, which in turn depends on the
geometry of the wavefront at the moment of the shock delivery. The lower
row of Figure 6 shows the case when, after the shock, the propagation resumes
not along the whole front, but only at the most concave segment of it,
where the front curvature assist the propagation. This is sufficient to
resume the rotation of the spiral wave. So, from this example it can be
seen that DFT measured in two dimensions should be usually higher than
that in one dimension.
We have calculated the one-dimensional DFT based on the properties of
the single cell version of the ventricular guinea-pig cell equations and
the restitution curve of original 1D model; this was found to be about
840nA/cell. The numerically computed 1D DFT was approx. 740 nA/cell, and
in 2D, approx. 750nA/cell. These values are for the rectangular current
pulses of 2 ms duration, and with the intracellular conductance assumed
10 micro S , which is, e.g., the conductance of a 30 micro m cube
of myoplasm with specific resistivity of 300 Ohm-cms . Assuming the orders
of magnitude for cell length, cell cross-section and heart cross-section,
an external current of 1000nA/cell corresponds to the electric field of
about 10V/cm and transcardiac current of 10A which quite agrees with the
experimental DFT of 5V/cm for electric field and 10 A for transcardiac
current; as we mentioned above, the theory allows absolute comparison with
experiment only in the order of magnitude. The close coincidence of 1D
and 2D estimations of DFT shows that the 2D effects are less important
than other simplifications used. We believe that the crudest of the simplifications
of that theory, after assumptions of uniformity of external current and
tissue properties, is the use of the Fife technique , considering the excitation
wave propagation as trigger waves in bistable media with one fast variable
(the transmembrane voltage), while the conditions of propagation are governed
by slow and local evolutions. The evolution in the OGPV model is more complicated,
as there are three other variables of characteristic time scales roughly
comparable to that of the transmembrane voltage.
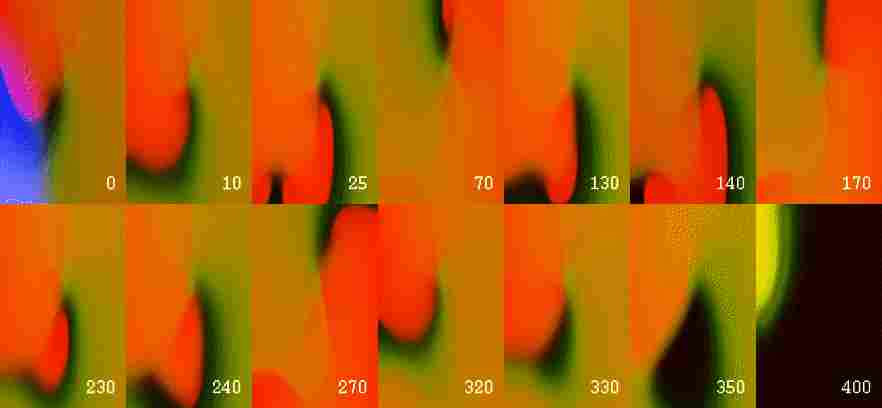
Figure 7: Elimination of a reentrant wave by a ``virtual electrode''
induced by stimulation of a near-DFT magnitude in full bidomain GPV model.
Top: first frame shows the area of the virtual electrode; other frames
show distribution of transmembrane voltage at selected time moments. The
time is in ms since the beginning of the stimulus.
3-dimensional aspects of re-entry in experimental and numerical models
of ventricular fibrillation
From Professor Jalife's laboratory at SUNY, Syracuse, we obtained experimental
visualisations of electrical activity from the endo- and epicardial surfaces
of pieces of sheep ventricular wall (5-11mm thick) that had been excised
and perfused via the coronary arteries, and superfused with oxygenated
physiological saline containing a drug (diacetyl monoxime), that blocked
contraction, and a potential-sensitive dye (di-4-ANEPPS). The video images
were obtained at 120 frames/s with a spatial resolution of approximately
0.5mm. The optical signals at different points were normalised to allow
for the variations of the dye concentration etc.

Figure 8: Delay coloured snapshots of surface views of experimental
polymorphic tachycardia in islated perfused wall of sheep ventricle. Top
row epicardial view, bottom row endocardial, with the interval between
images 50 ms.
The typical qualitative properties of experimentally observed excitation
patterns can be summarised as follows.
-
Synchronous endo- and epicardial views of the same preparation can, and
most often do, show different dynamics. In case of simple excitation pattern,
corresponding to monomorphic tachycardia, the patterns are different but
synchronous; in more complex cases, corresponding to polymorphic tachycardia/fibrillation,
they seem virtually independent.
-
At every particular point, most of the time the electrical activity is
approximately periodic. The spatio-temporal pattern as a whole can be approximately
periodic, in the examples that correspond to monomorphic tachycardia, but
not in the examples that correspond to polymorphic tachycardia/fibrillation.
-
During fibrillation, spiral waves are sometimes seen on the surfaces, but
quite often they are not. If they are seen, they appear only transiently,
for a few rotations, and then disappear.
-
The (visual) complexity of the patterns changes with time; at large times,
it appears to increase.
All these observations are consistent with scroll waves of excitation within
the bulk of the ventricular wall [41].
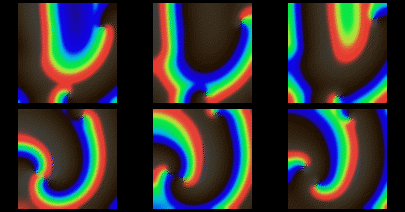
Figure 9: Numerical solutions of qualitative features of surface
views of polymorphic tachycardia seen in Figure 8, using FitzHugh-Nagumo
model in a 50 s.u cube, time interval between images 4.2 t.u.
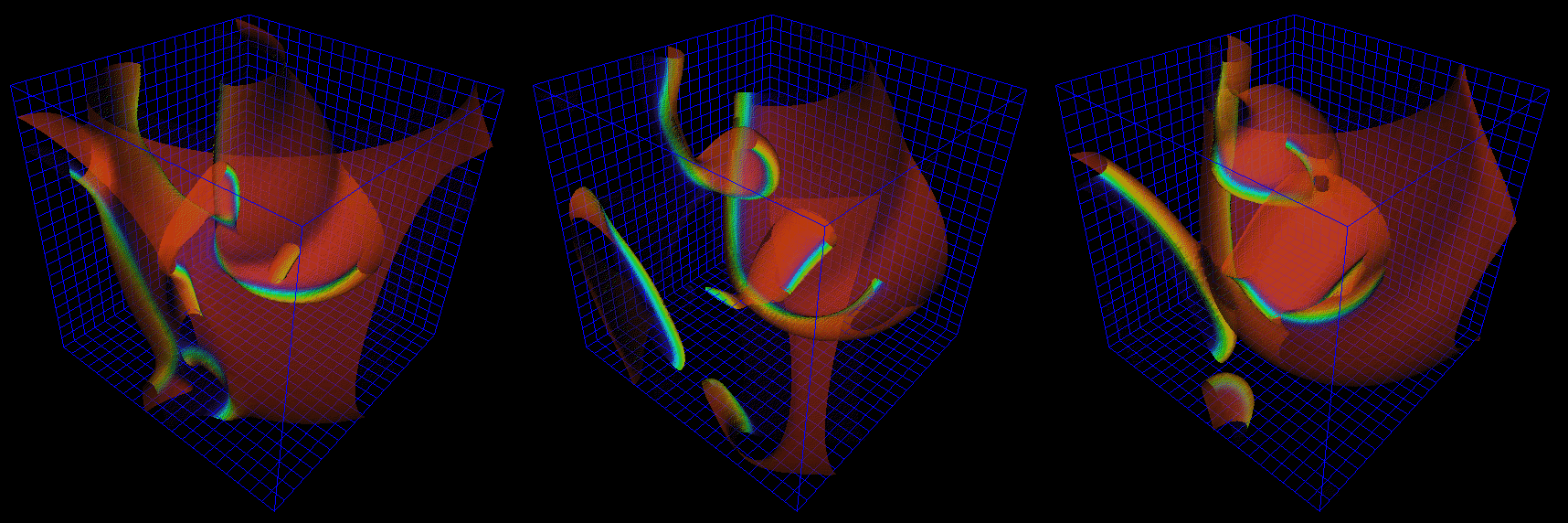
Figure 10: Numerical solutions of qualitative features of three-dimensional
mechanism generating polymorphic tachycardia simulation seen in Figure
8, using FitzHugh-Nagumo model in a 50 s.u cube, time interval between
images 4.2 t.u.
The choice of parameters used in the simulations of Figures 9 and 10
provides a negative tension of the filaments, i.e. scroll waves
in sufficiently large media are unstable, their filaments tend to lengthen,
curve, touch the boundaries and each other and break onto pieces, each
of which then grows again [42] etc. With the same parameter
values, the same set of equations in two spatial dimensions shows quite
stable spiral waves. This is in qualitative correspondence with the fact
that real fibrillation is only observed in sufficiently thick hearts or
heart preparations.
The differences in spatial activity on the two surfaces demonstrate
the essentially three-dimensional nature of the electrical activity that
generates fibrillation in the animal tissue model. The computations show
that the patterns of activity can, in principle, be accounted for by scroll
waves within the ventricular wall. The scroll waves used to reproduce the
surface patterns are roughly parallel to the ventricular surfaces, in contrast
to the transmural filament proposed in [43]. In an
intact heart, these waves would be around filaments which are closed (
i.e. scroll rings) or that terminate an inexcitable boundary.
Domain structure during ventricular fibrillation
Quantitative analysis of the excitation pattern on the cardiac surfaces
has lead to the observation that the dominant frequency of oscillations
has a domain structure, the frequency being approximately uniform within
any one domain , and the boundaries between the domains are being sharp
(of the order of 1 mm), and the domains persist over minutes [44].
This reconciles the contradiction between the recent description of order
in fibrillation, based on statistical analysis of high-resolution data,
with the traditional picture of disordered fibrillation based on low-resolution
maps, single electrograms or ECG.
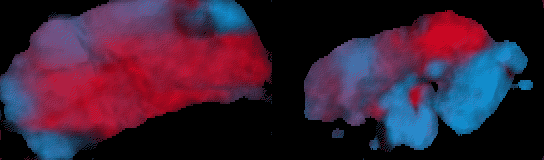
Figure 11: Experimental data illustrating the frequency domains.
Blue and red components of the painting show the power of the two frequency
components at each point; the spatial separation of the colours is the
demonstration of the domain structure of the excitation pattern. The ratio
of frequencies is 15 : 12.5 = 6 : 5.
These domains could be due to different re-entrant sources with different
periods, or could be produced by one re-entrant vortex with a period shorter
than the minimal propagation period of some parts of the tissue, and the
domains could be produced by frequency division due to partial conduction
block. This presupposes heterogeneity in the tissue properties. Although
it is easy to distinguish between these mechanisms using data generated
in simulations, by constructing power spectra (where the ratio of dominant
frequencies will be ratio of small integers), the frequency broadening
due to the short duration of episodes of fibrillation means that frequency
ratios cannot provide a practical tool for distinguishing between the two
methods. However, when combined with Lissajous figures the experimental
records can be separated into those in which the frequency patterns are
consistent with conduction block and those in which several re-entrant
sources cannot be excluded [45 46]

Figure 12: Numerical solutions illustrating the frequency domains.
Blue and red components of the painting show the power of the two frequency
components at each point; the spatial separation of the colours is the
demonstration of the domain structure of the excitation pattern. All dynamical
variables in the right half of the medium have been slowed, and there is
a single spiral source that is pinned in the left half of the medium.
Conclusions
The models for cell excitation which are incorporated into the virtual
tissues are based on extensive, in vitro experiments and so they
have a firm experimental basis. The key assumption in the virtual tissues
we have presented is that propagation phenomena can be represented by a
spatially continuous, rather than discrete, cell-to-cell process. If this
assumption is valid then the phenomenology presented should be seen in
tissue experiments, and optical recordings of electrical activity on the
heart surface are begining to provide an experimental basis that can be
used to validate the applicability of the virtual tissue behaviours.
It is now technically feasible to incorporate the virtual tissue models
into anatomically realistic geometry and fibre orientation, and to incorporate
transmural and regional differences of excitation processes. Thus, a virtual
organ (the ventricle) can be used to explore the mechanisms of propagation
disorders. The incorporation of excitation-contraction coupling is well
under way, and the ability to interact, using haptic feedback and tissue
mechanics, with such an electro-mechanical virtual organ is under development.
References
[1] Boyettt, M.R., Clough, A., Dekanski, J. and Holden,
A.V. " Modelling Cardiac Excitation and Excitability" in Panfilov, A.V.
and Holden, A.V., eds., The Computational Biology of the Heart, Wiley:Chichester,
ISBN 0-471-96200-9, 1997.
[2] Biktashev, V.N. and Holden, A.V., "Re-entrant
activity and its control in a model of mammalian ventricular tissue", Proceedings
of the Royal Society (London), vol B263, pp 1373-1382, 1996.
[3] Zhang, H, Holden, A.V., Kodama, I.M Honjo, H.,
Lei, M, Varghese, T. and Boyett, M.R., "Mathematical models of action potentials
in the periphery and center of the rabbit sinoatrial node", American Journal
of Physiology (Heart and Circulatory Physiology), vol. 279, in press, 2000.
[4] Noble, D., "Oxsoft HEART Version 3.8 Manual",
Oxsoft: Oxford, 1990.
[5] Bower, J.D and Beeman, D., "The Book of GENESIS",
Springer-Verlag: New York, ISBN 0-387-94019-7, 1994.
[6] Zhang, H, Holden, A.V. and Boyett, M.R., "Computer
modelling of the sinoatrial node", this volume, 2000.
[7] Li, L., Zhang, H, Holden, A.V. and Orchard, C.H.,
"Computer modelling of the sinoatrial node", this volume, 2000.
[8] Spach, M.S., "Microscopic Basis of Anistropic
Propagation in the Heart and the Nature of Current Flow at a Cellular Level"
in Zipes,D.P. and Jalife, J., eds., Cardiac Electrophysiology - from cell
to bedside, Second Edition , W.B.Saunders, Philadelphia, ISBN 0-7216-4941-6,
1995.
[9] Winslow, R.L., Kimball, A., Varghese, T. and
Noble, D., "Simulating cardiac sinus and atrial network dynamics on the
Connection Machine", Physica-D, vol. 64, pp 281-298, 1993.
[10] Keener, J.P., "The effects of discrete gap
junction coupling on propagation in myocardium", Journal of Theoretical
Biology, vol. 148, pp 49-82, 1991.
[11] Hunter, P.J., Smaill, B.H., Nielsen, P.M.F.,
amd Le Grice, I.J., "A mathematical model of cardiac anatomy" in Panfilov,
A.V. and Holden, A.V., eds., The Computational Biology of the Heart,Wiley:Chichester,
ISBN 0-471-96200-9, 1997.
[12] Panfilov, A.V. and Keener, J.P., "Re-entry
in three-dimensional FitzHugh-Nagumo medium with rotational anisotropy",
Physica-D, vol. 84, pp 545-552, 1995.
[13] Panfilov, A.V., "A mathematical model of cardiac
anatomy" in Panfilov, A.V. and Holden, A.V., eds., The Computational Biology
of the Heart, Wiley:Chichester,ISBN 0-471-96200-9, 1997.
[14] Holden, A.V., Poole, M.J. and Tucker, J.V.,
"An Algorithmic Model of the Mammalian Heart: Propagation, Vulnerability,
Re-entry and Fibrillation", International Journal of Bifurcation and Chaos,
vol. 6, pp 1623-1636, 1996.
[15] Biktashev, V.N., Biktasheva, I.V., Brindley,
J., Holden, A.V., Hill, N.A. and Tsyganov, M.A., "Effects of shear flows
on nonlinear waves in excitable media ",Journal of Biological Physics,
vol.25, pp 101-113, 1999.
[16] Biktashev, V.N., Holden, A.V., Tsyganov, M.A.,
Brindley, J. and Hill, N.A., "Excitation wave breaking in excitable media
with linear shear flow", Physical Review Letters , vol.81, pp 2815-2818,
1998.
[17] Roth, B.J. and Krassowska, W. , "The induction
of reentry in cardiac tissue. The missing link: How electric fields alter
transmembrane potential", Chaos , vol.8, pp. 204-220, 1998.
[18] Trayanova, N., Skuibine, K. and Aguel, F. ,
"The role of cardiac tissue structure in defibrillation", Chaos , vol.8,
pp, 221-233, 1998.
[19] Sobie, E.A., Susil, R.C. and Tung, L., "A generalized
activating function for predicting virtual electrodes in cardiac tissue",
Biophysical Journal , vol.73, pp. 1410-1423, 1997.
[20] Keener, J.P. and Bogar, "A numerical method
for the solution of bidomain equations in cardiac tissue", Chaos , vol.8,
pp 234-241, 1998.
[21] Biktashev, V.N., Holden. A.V. and Zhang, H.,
"A model for the action of external current onto excitable tissue", International
Journal of Bifurcation and Chaos , vol.7, pp 477-485 , 1997.
[22] Starmer, C.F., Biktashev, V.N., Romashko, M.R.,
Stepanov, M.R., Makarova, O.N. and Krinsky, V.I., "Vulnerability in an
excitanble medium: Analytical and Numerical Studies of Initiating Unidirectional
Propagation", Biophysical Journal vol.65 pp.1775-1787, 1993.
[23] Cardiac Arrhythmia Suppression Trial (CAST)
Investigators, "Preliminary report: effect of encainide and flecainide
on mortality in a randomised trial of arryhythmia suppression after myocardial
infarction", New England Journal of Medicine, vol. 321 pp.406- ,1989.
[24] Biktashev,V.N., Holden, A.V. and Nikolaev,
E.V., "Spiral wave meander and symmetry of the plane", International Journal
of Bifurcation & Chaos, vol 6, pp 2433-2440, 1996.
[25] Nikolaev, E.V., Biktashev, V.N. and Holden.,
A.V. "On bifurcations of spiral waves in the plane", International Journal
of Bifurcation and Chaos, vol. 9, pp. 1501-1516, 1999.
[26] Holden:, A.V., "The restless heart of a spiral",
Nature, vol. 387 pp. 655-6, 1997.
[27] Biktashev, V.N. and Holden. A.V., "Control
of re-entrant activity in a model of mammalian atrial tissue", Proceedings
of the Royal Society (London) B, vol 260, pp. 211-217, 1995.
[28] Holden, A.V. and Zhang, H., " Charactertistics
of atrial re-entry and meander computed from a model of a single atrial
cell", Journal of Theoretical Biology 1995
[29] Clayton, R.H., Murray, A., Higham, P.D. and
Campbell, R.W.F. "Self-terminating ventricular tachyarrhythmias - a diagnostic
dilemma", Lancet vol. 341, pp.93-95, 1993.
[30] Zareba, W., Moss, A.J., Schwartz, P.J. et al.,
"Influence of the genotype on the clinical course of the long QT syndrome",
New England Journal of Medicine vol. 339 pp.960-965, 1998.
[31] Ackermann, M.J. and Clappham, D.E., "Ion channels
- basic science and clinical disease", New England Jonal of Medicnie vol
336 pp1575-86, 1997
[32] Roden, D.M. and Balser, J.R., "A plethora of
mechanisms in the HERG related long QT syndrome - genetics meets electrophysiology",
Cardiovascular Research vol. 44, pp. 242-246, 1999.
[33] Clayton, R.H., Bailey, A., Biktashev, V.N.
and Holden, A.V.,"Re-entrant arrhythmias in simulations of the Long-QT
syndrome", Computers in Cardiology vol. 26, pp.121-124, 1999.
[34] Biktashev, V.N. and Holden, A.V., "Resonant
drift of autowave vortices in two dimensions and the effects of boundaries
and inhomogeneities", Chaos Solitons and Fractals vol. 5 pp. 575-622, 1995.
[35] Biktashev, V.N. and Holden, A.V., "Design principles
of a low voltage cardiac defibrillator based on the effect of feedback
resonant drift", Journal of Theoretical Biology vol.169 pp 101-112, 1994.
[36] Lab, M.J., "Fibrillation, chaos and clinical
control", Nature Medicine vol. 3 pp. 385-6 1997.
[37] Nikolaev, E.V., Biktashev, V.N. and Holden,
A.V., "On feedback resonant drift and interaction with the boundaries in
circular and annular excitable media", Chaos Solitons and Fractals, vol.
8, pp363-377, 1998.
[38] Elkin, Yu.E., Biktashev, V.N. and Holden, A.V.,
"On the movement of excitation wavebreaks", Chaos Solitons and Fractals
vol.9 pp.1597-1610 , 1998.
[39] Winfree, A.T, "Varieties of spiral wave behaviour:
an experimentalist's approach to the theory of excitable media " Chaos
vol.1, pp.303-334m 1991.
[40] Biktashev, V.N. and Holden,A.V., " Deterministic
Brownian motion in the hypermeander of spiral waves " Physica-D vol. 116
pp 342-354, 1998.
[41] Biktashev, V.N., Holden, A.V., Mironov, S.F.,
Pertsov, A,M. and Zaitsev, A.V., " Three dimensional aspects of re-entry
in experimental and numerical models of ventricular fibrillation. " International
Journal of Bifurcation and Chaos, vol9, pp. 695-704, 1998.
[42] Biktashev, V.N., Holden, A.V. and Zhang, H.,
"Tension of organizing filaments of scroll waves", Philosophical transactions
of the Royal society of London: Physical Sciences and Engineering, vol.
347 pp.611-630, 1994.
[43] Fenton, F. and Karma, A. "Vortex dynamics in
three-dimensional continuous myocardium with fiber rotation: Filament instability
and fibrillation", Chaos, vol. 8, pp.20-47, 1999.
[44] Zaitsev, A.V., Berenfeld, O, Mironov,S.F.,
Jalife.J. and Pertsov, A.M., "Distribution of excitation frequencies on
the epicardial and endocardial surfaces of fibrillating ventricular wall
of the sheep heart", Circulation Research, vol.86, pp.408-417 2000.
[45] Biktashev, V.N., Holden, A.V., Mironov, S.F.,
Pertsov, A.M. and Zaitsev, A.V., "On the mechanism of the domain structure
of ventricular fibrillation - a case study", International Journal of Bifurcation
and Chaos, in press 2000.
[46] Biktashev, V.N., Holden, A.V., Mironov, S.F.,
Pertsov, A.M. and Zaitsev, A.V., "Two mechanisms of the domain structure
of ventricular fibrillation", Journal of Physiology in press 2000.
|

 Home
Current Issue
Table of Contents
Home
Current Issue
Table of Contents